Introduction
Over the last century, the Philippines has significantly lost its forest. This decline was brought about by massive deforestation through extensive logging. To combat this problem, large-scale reforestation efforts have been implemented by both the government and the private sector (Tolentino, 2008). Unfortunately, of ten most planted trees in reforestation projects around the country, eight are exotic species, with large leaf mahogany (Swietenia macrophylla ) being the most dominant (Tolentino, 2008). Furthermore, 80% to 90% of trees planted under the Philippine government’s National Greening Program are exotic (Torres, 2018).
In the Philippines, exotic species introductions might have begun during the pre-historic period because of the Indo-Malayan origin of common agricultural tree crops (Baguinon et al ., 2005). Alien or exotic species, whether intentionally or unintentionally introduced into a new habitat, significantly can alter the native structure and function of an area. These fast-growing exotics are preferred over native trees because of their high economic value. However, they have the potential to alter ecosystem functions and soil attributes (Tulod et al ., 2017). Bioinvasion happens when exotic species proliferate, persist, and dominate distant ranges, thus suppressing native species (Florece & Baguinon, 2011; Hejda et al ., 2009). Moreover, intercontinental commerce and trade are major drivers in the spread and local colonization of exotic species (Florece & Baguinon, 2011). These species also grow very fast with high fruit yields. In addition, they can tolerate stress in marginal sites (Tolentino, 2008).
Exotic to the Philippines, large leaf mahogany (Swietenia macrophylla King) is a fast-growing, deciduous, canopy emergent tree native to the seasonally dry tropical forests of Mexico down to Bolivia (Grogan et al ., 2014). It is the world’s most valuable and widely traded tropical timber species. It is being exploited since the Spanish discovery in the 16th century due to its unique properties (Mejia et al ., 2008). It was first planted in Manila, Philippines in 1907, then at Mt. Makiling, Laguna in 1913 (Baguinon et al ., 2005). Until 2011, it was the most planted tree in the Philippine government’s National Greening Program (Torres, 2018). It is regarded as bioinvasive in the Philippines (Baguinon et al ., 2005). Castillo (2001) has reckoned that S. macrophylla has a negative effect on species diversity and evenness of native taxa, indicating successful bioinvasion of S. macrophylla . Moreover, Thinley’s (2002) laboratory experiments have shown that S. macrophylla leaf extract can significantly inhibit the growth of Pterocarpus indicus (a Philippine native tree), having the greatest negative effect at the juvenile stage. In addition, S. macrophylla leaves contain allelochemicals including moderate amounts of phenols, appreciable amounts of tannins, and substantial amounts of alkaloids. These results support the widely accepted notion that S. macrophylla has allelopathic effects on neighboring plant taxa.
Bioinvasion as an environmental threat has long been recognized by scientists since the 1950s (Florece & Baguinon, 2011). While other countries have made significant efforts in understanding impacts of exotic invasive species to ecosystems, the Philippines has just recently acknowledged this phenomenon. Little is known about the roles of exotic species in natural ecosystems (Tulod et al ., 2017). Thus, the aim of this study was to determine effects of large leaf mahogany (Swietenia macrophylla ) on the growth and survival of a native Philippine tree species naturally co-occurring with large leaf mahogany. Specifically, we sought to: (1) evaluate the in situ effect of large leaf mahogany leaf litter on the growth of native Pterocarpus indicus seedlings; and (2) determine the effect of large leaf mahogany canopy on the growth of P. indicus seedlings. We conducted in situ experiments to evaluate the effect of large leaf mahogany on natural ecosystems.
Materials and Methods
Test taxon
Pterocarpus indicus was chosen as the test taxon for this study mainly due to its natural co-occurrence with S. macrophylla and its abundance at study sites. Pterocarpus indicus (family Fabaceae) is a pioneer tree species native to the Philippines and widely valued for its hardwood. Pterocarpus indicus seeds were collected in September 2018. These seeds were surface sterilized with 70% (v/v) ethanol for 60 seconds and 4% (v/v) sodium hypochlorite for 45 seconds. They were then rinsed six times with steriledistilled water (Lok et al ., 2006) and sown in Petri dishes. After one week, germinants were transplanted to plastic pots and grown for five months.
Effects of S. macrophylla leaf litter and canopy
Three sites dominated by S. macrophylla trees inside the University of the Philippines, Diliman campus, were selected. A pair of 10 x 10m2 plots were established in each of these three sites. Twenty P. indicus seedlings were introduced and planted in each plot of 10 x 10m2 For each plot pair, leaf litter in one plot was removed every other day while no leaf litter removal was done for the other plot. Growth reported as root collar-to-shoot heights of all P. indicus seedlings were recorded every five days for one month. Percent canopy of each plot was also recorded using a densiometer (Forestry Suppliers, Inc., USA) to determine the effect of S. macrophylla canopy closure on P. indicus seedling growth.
To have baseline information about the growth of P. indicus seedlings, we established a control plot in a plant box without having contact with a single S. macrophylla tree. We also planted 20 P. indicus seedlings inside this control plot.
Data analyses
Root collar-to-shoot height was reported as standardized growth rate. It was calculated by subtracting the initial height of P. indicus seedling from its final height divided by the initial height. The standardized growth rate was binomially distributed. Therefore, it was analyzed using a Generalized Linear Model with quasi-binomial errors to account for overdispersion. The presence of S. macrophylla leaf litter, average percent canopy closure, and frequency of S. macrophylla trees per study site were included as independent variables. Standardized growth rate was used as a dependent variable. All analyses were performed in R ver. 3.6.3 (R Core Team, 2020).
Results
Effect of S. macrophylla leaf litter
Smoothed plots (Fig. 1) showing heights of P. indicus seedlings in the first day as compared to those in succeeding days of observation indicated that after one month, seedlings in plots without S. macrophylla leaf litter (Absent panel) grew faster than seedlings in plots with leaf litter (Present panel). In addition, the control plot tended to follow the pattern of plots without leaf litter (Fig. 1).
Fig. 1.
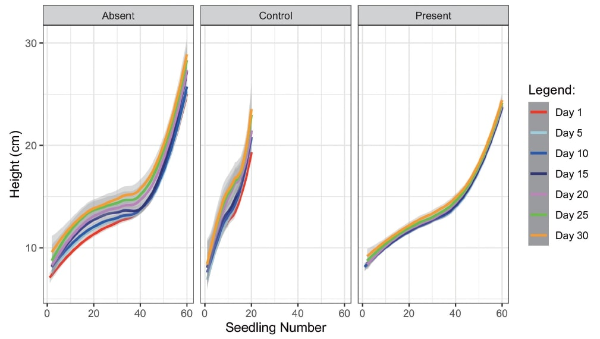
The standardized growth rate of seedlings in plots without leaf litter was significantly higher than that of seedlings in plots with leaf litter (Fig. 2, GLM with quasi-binomial errors: p <0.0001).there was no significant difference between the standardized growth rate of the seedlings in plots without leaf litter and that of seedlings in the control plot (Fig. 2, GLM with quasi-binomial errors: p = 0.329).
Fig. 2.
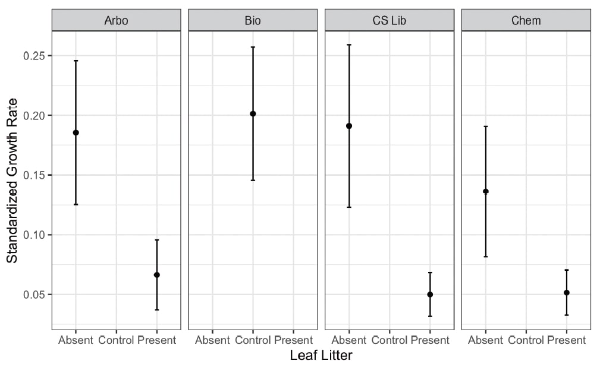
Effect of S. macrophylla frequency in study sites
The abundance of S. macrophylla trees per study site (Table 1) was examined to determine its effect on the mean standardized growth rate of seedlings between plots with and without leaf litter (Fig. 2). Results showed that S. macrophylla tree abundance did not significantly affect the standardized growth rate of P. indicus seedlings (GLM with quasi-binomial errors: p = 0.162).
Table 1.
| Study Site | S. macrophylla tree frequency | S. macrophylla leaf litter presence | Average % canopy closure |
|---|---|---|---|
| Site 1 (Arbo) | 20 | Present | 75.93 |
| Absent | 78.23 | ||
| Site 2 (Bio) | 0 | Absent | 37.43 |
| Present | 76.34 | ||
| Site 3 (CS lib) | 35 | Absent | 72.91 |
| Present | 63.22 | ||
| Site 4 (Chem) | 8 | Absent | 59.03 |
Effect of S. macrophylla canopy
Average percent canopy closure (Table 1) was also examined to determine its effect on the standardized growth rate. Results showed that large leaf mahogany canopy did not significantly affect the height of P. indicus seedlings in all plots (GLM with quasi-binomial errors, p = 0.322).
Discussion
With the staggering loss of forest cover in the Philippines starting from the Spanish colonization and peaking during the American period, forest rehabilitation is critically important for the conservation of its biodiversity resources. Threats to its remaining forests is further exacerbated by human activities (Malabrigo et al ., 2016). Considering the amazing diversity of native trees in the Philippines, species used in forest restoration are surprisingly few. In these initiatives, 95% are exotic species (Malabrigo et al ., 2016). Restoration programs have focused on the use of exotic trees owing to a misconception about native species’ silvicultural requirements and the lack of management information (Butterfield, 1995; Chechina & Hamann, 2015). Furthermore, exotic trees are believed to be able to regain forest cover faster, stabilize watersheds, and reduce downstream flooding (Torres, 2018).
Deleterious effects of Swietenia macrophylla on Philippine native trees
widely planted exotic tree species Swietenia macrophylla on a valuable Philippine native tree species Pterocarpus indicus. Specifically, the in situ growth of P. indicus seedlings was significantly inhibited by the leaf litter of S. macrophylla . Thinley (2002) has also found that S. macrophylla leaf extract as watering medium can inhibit the growth of P. indicus due to its leaf allelochemicals (moderate amounts of phenols, appreciable amounts of tannins, and substantial amounts of alkaloids). Mukaromah et al . (2017) have also suggested the possibility of the allelopathic potential of S. macrophylla by showing a concomitant inhibition of radicle lettuce seedling growth with increasing S. macrophylla leaf litter concentration. Taken together, these findings indicate that S. macrophylla leaves can negatively affect seedling growth of P. indicus both in situ and ex situ.
The presence of S. macrophylla also seems to negatively affect native tree biodiversity. In our study area, there were 184 native tree species. However, only 33 (17.93%) were found to coexist with S. macrophylla. Fig. 3A shows an area with little undergrowth dominated by S. macrophylla . In contrast, Fig. 3B shows an area dominated by indigenous trees where the undergrowth plant community is thriving. Moreover, most trees that coexist with S. macrophylla had basal area of less than 50cm2 (diameter = 15 cm). This might imply that even though they survive and coexisted with S. macrophylla , their growth might be inhibited. In some of these species, individuals that were not close to S. macrophylla trees had larger basal areas (e.g., Pterocarpus indicus , Dracontomelon dao , and Terminalia catappa , up to 300cm2; Koordersiodendron pinnatum, Dysoxylum gaudichaudianum, Calophyllum inophyllum, Reutealis trisperma and Premna odorata , up to 100cm2 Baguinon et al . (2008) have also shown that forest diversity decreases when the importance value of S. macrophylla increases, suggesting a competitive exclusion against native trees. This might be due to the thick shade and leaf shedding characteristics of S. macrophylla that can prevent the recruitment of native species. The dry environment provided by the thick leaf litter mat shed by S. macrophylla can cause germination failure in recalcitrant seeds of native species (Baguinon & Miel, 2012).
Fig. 3.

Moving forward with restoring our forests
Based on overwhelming scientific evidence showing the negative effect of S. macrophylla on native trees, we highly recommend its discontinued use in tree planting and reforestation efforts. Unfortunately, correcting mistakes of the past will be an uphill battle. Swietenia macrophylla grows in tropical areas in a wide variety of climatic and edaphic conditions (Navarro-Martínez et al ., 2018). It is found in all forest types, mostly in hardwood forests, along riverbanks, and on deep alluvial soils (Mejia et al ., 2008). Furthermore, it is a pioneer species that bears capsules with winged seeds that can be dispersed 20 to 40 meters from the mother tree (Baguinon et al ., 2005; Brown et al ., 2003). Each capsule contains about 60 recalcitrant seeds that can germinate in less than a month. These seeds contain food reserves and undergo hypogeal germination. Thus, young S. macrophylla can develop even without an initial photosynthesis in poorly lit areas. Moreover, its leaves are rarely attacked by herbivores (Baguinon et al ., 2005).
Faced with this current situation in the Philippines, we recommend a two-pronged approach: (1) active management of S. macrophylla -planted reforestation sites; and (2) widespread use of native species. Active management is needed because exotic species can negatively affect ecosystem functions, making restoration sites inhospitable for native species. These efforts may include leaf-litter removal as S. macrophylla trees shed their leaves every February (Baguinon et al ., 2005), branch pruning, and gradual removal of S. macrophylla trees. Our results point to the importance of leaf-litter removal. However, more studies are needed to elucidate the best approach to actively manage S. macrophylla trees.
In recent years, different initiatives have started to use native trees in reforestation efforts because of their importance in restoring forest cover and ecosystem services (Malabrigo et al ., 2016; Tolentino, 2008). Tulod et al . (2017) have noted that native forests are more favorable habitats for local biodiversity than exotic forests as evidenced by high species richness and diversity in native forests compared to exotic tree plantations. Thus, the use of native trees in forest restoration efforts should be of the utmost priority. This will ensure a balanced ecosystem dominated by native flora and fauna.
Acknowledgments
We sincerely thank Ramon Bandong, Arthur Glenn Umali and Dr. Bonifacio Pasion for help in identifying uncommon species and Dr. Teresita Perez for comments on the earlier version of this manuscript. We would also like to acknowledge the UP Diliman College of Science for providing permits to conduct field experiments at our study sites.
References
(2001) Vegetation analysis of undergrowth plants in lowland forest of Mt. Makiling as a tool in assessing the advance and spread of big leaf mahogany (Swietenia macrophylla King ) (Unpublished B.S. Forestry thesis). University of the Philippines Los Banos-College of Forestry and Natural Resources Library, Laguna, Philippines. .
, , , (2008, Retrieved April 14, 2020) Big-leaf mahogany (Swietenia macrophylla ) in Peru, Bolivia and Brazil .Case study for: International Expert Workshop on CITES Non-Detriment Findings. from http://www.conabio.gob. mx/institucion/cooperacion_internacional/TallerNDF/ Links-Documentos/WG-CS/WG1-Trees/WG1-CS4%20 Swietenia/WG1-CS4.pdf
(2002) Unpublished B.S. Forestry thesis Laguna, Philippines.: University of the Philippines Los Banos-College of Forestry and Natural Resources Negative interaction between large leaf Mahogany (Swietenia macrophylla) and some Indigenous tree species in lowland forest of Mt. Makiling- Allelopathy, a possible cause?