Introduction
Wetlands are transitional zones between terrestrial and aquatic ecosystems (Yi & Nam, 2008) that are permanently or seasonally humid (Park et al ., 2000). They are characterized by high productivity and rich biodiversity. Wetlands provide humans with various ecological ser vices by offering an abundance of natural resources and creating unique natural landscapes (Ahn et al ., 2016 ; Kim 2003). Depending on where they are formed, wetlands can be categorized as coastal wetlands, inland wetlands (riverine, lacustrine, and mountainous palustrine), or constructed wetlands, each of which has its value as a unique ecological habitat (Ministry of Environment (ME),2010).
The Civilian Control Zone (CCZ) in South Korea is located 5-20 km south of the demilitarized zone (DMZ), consisting of 15 cities or counties (si/gun>) and 98 eup/myeon (Lee, 2004). Civilian access to this region has been restricted for approximately 60 years, leading to the formation of a unique natural ecosystem that cannot be found in other regions (Cha et al ., 2000). There is a broad distribution of wetlands and paddy fields that display high biodiversity and ecosystem connectivity (Park & Nam, 2013). The entire region outside the DMZ, including the CCZ, consists of 3.3% of wetlands (Lim, 2006). Yongyangbo Wetlands, located close to the southern border of the DMZ, consist of lacustrine wetlands formed owing to the influence of Yongyang Weir and the riverine wetlands between Yongyang Weir and Sinamjeongyo Bridge. The Hwagang River, which is a branch of the Hantangang River, originates from Suribong Peak, Geumseong myeon, Kimhwa gun, entering South Korea after it crosses the Military Demarcation Line. Sediment carried by the river accumulates upstream of Yongyang Weir, which is believed to have led to the formation of lacustrine wetlands (National Institute of Ecology (NIE), 2019).
Several survey studies on forests have focused on the distribution of plants throughout the DMZ, starting with a study on the distribution of Pinus densiflora Siebold & Zucc. (Lee, 1968; Moon, 2019). Recent studies have been conducted in Gwangchiryeong, Yang-gu (Heo et al ., 2017) and Daedeukbong Peak, Cheolwon gun (An et al ., 2018). In terms of wetland studies, some studies have been conducted on the flora in the Hak Reservoir, which is connected to the Hantangang River water system in the region bordering the DMZ (Shin et al ., 2015)
In this study, we performed a floristic survey of Yongyangbo Wetlands in the CCZ under circumstances that ecological surveys, including floristic surveys, were limited by restricted access to the region. This is necessary for military reasons. It is also necessary due to local characteristics such as the presence of mines scattered around the area (Oh et al ., 2009). Through this survey, we ascertained the distribution of endangered wild plant habitats and the distribution of naturalized plants that might threaten the ecosystem. Results of our survey can be used as basic data to support the conservation of wetland ecosystems in the CCZ and for the management of Yongyangbo Wetlands, which have been designated as wetland protected areas.
Materials and Methods
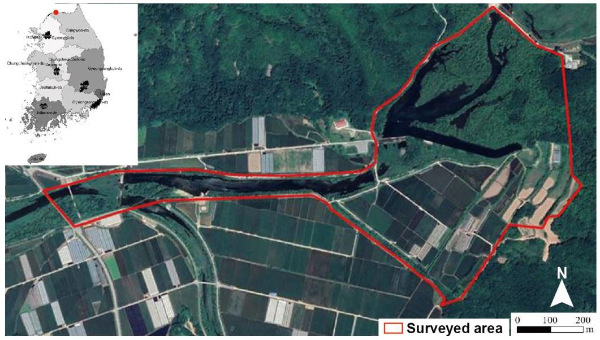
Survey area
In terms of administrative districts, Yongyangbo Wetlands bridge Yongyang-ri and Amjeong-ri in Kimhwa-eup, Cheolwon-gun, Gangwon-do, and South Korea. Specifically, it is a region between 37°12′08″N and 37°14′00″N, 127°42′48″E, and 127°44′22″ E (Fig. 1). The survey area was about 0.77 km2 with an average altitude of 227.7m and an average slope of 2.8°. According to data from the Cheolwon Weather Station, over the last 30 years (1989-2018), the mean annual air tempera ture was 10.25°C and the mean annual precipitation was 1,381.14 mm in the Yongyangbo Wetlands as a whole, which was similar to the mean annual precipitation across South Korea. Approximately 72.5% of the total annual precipitation occurs in summer (June to September), showing typical rainfall characteristics of a temperate monsoon region, with large differences in precipitation between seasons (NIE, 2019).
Survey method
A field survey of five sessions was conducted between April and August 2019 in accordance with the National Wetland Survey Protocol (ME, 2011). The survey was conducted on foot, covering the entire Yongyangbo Wetlands. Identification and classification wer e performed as described by Lee (1996a), Lee (1999), and Park (2009). Scientific and Korean names were assigned in accordance with the National List of Species of Korea (National Institute of Biological Resources (NIBR), 2020). Plant lists were grouped by taxonomic ranks of family and below and arranged in alphabetical order. Based on plants identified in the survey, life forms (Lee, 199 6b), endemic plants (Chung et al ., 2017), floristic target species (NIE, 2018), and rare plants (Korea National Arboretum (KNA), 2008) were summarized. Naturalized plants were investigated as described by Lee et al . (2011) and Ryu et al . (2017). Floristic richness (Kim & Lee, 2006) was analyzed to investigate the biodiversity of plants in the survey. To ascertain the degree o f wetland disturbance, disturbance index (Benabdelmoumence et al ., 2014) and naturalization index (Numata, 1975) were calculated.
Results and Discussion
Flora of vascular plants
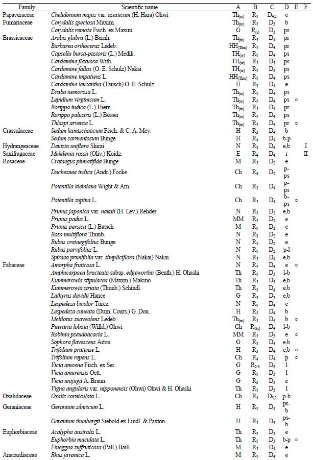
The flora observed at Yongyangbo Wetlands consisted of a total of 248 taxa, 72 families, 163 genera, 230 species, 4 subspecies, and 14 varieties. There were 4 taxa of Pteridophyta (4 families, 4 genera, 4 species), 1 taxon of Pinophyta (1 family, 1 genus, 1 species), and 243 taxa of Magnoliophyta (67 families, 158 genera, 164 species, 4 subspecies, 14 varieties; Table 1, Supplementary Table 1 ), corresponding to approximately 5.4% of 4,596 vascular plant taxa found in the Korean Peninsula (NIBR, 2020). The m ost commonly observed families in order of descending richness were Poaceae (26 taxa, 10.5%), Asteraceae (25 taxa, 10.1%), Cyperaceae (17 taxa, 6.9%), and Fabaceae (17 taxa, 6.9%; Table 2). The floristic diversity of Yongyangbo Wetlands was 1.52, higher th an that of Hak Reservoir near the DMZ in Cheolwon (1.38, Shin et al ., 2015) but lower than that of Daedeukbong Peak (1.59, An et al ., 2018). Floristic diversity was calculated as the species to genera ratio, with a larger value indicating more diverse genera containing a broader spectrum of species (Kim & Lee, 2006).
Table 1.
| Taxa | Fam. | Gen. | Sp. | Subsp. | Var. | For. | Total |
|---|---|---|---|---|---|---|---|
| Pteridophyta | 4 | 4 | 4 | - | - | - | 4 |
| (Polypodiopsida) | (3) | (3) | (3) | - | - | - | (3) |
| (Equisetopsida) | (1) | (1) | (1) | - | - | - | (1) |
| Pinophyta | 1 | 1 | 1 | - | - | - | 1 |
| (Pinopsida) | (1) | (1) | (1) | - | - | - | (1) |
| Magnoliophyta | 67 | 158 | 225 | 4 | 14 | - | 243 |
| (Magnoliopsida) | (55) | (124) | (164) | (4) | (13) | - | (181) |
| (Liliopsida) | (12) | (34) | (61) | - | (1) | - | (62) |
| Total | 72 | 163 | 230 | 4 | 14 | - | 248 |
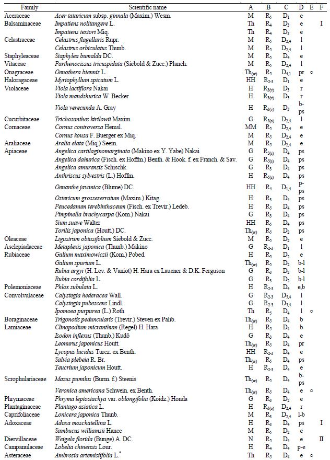
Table 2.
| Family | No. of taxa | Ratio (%) |
|---|---|---|
| Poaceae | 26 | 10.5 |
| Asteraceae | 25 | 10.1 |
| Cyperaceae | 17 | 6.9 |
| Fabaceae | 17 | 6.9 |
| Polygonaceae | 12 | 4.9 |
| Brassicaceae | 12 | 4.9 |
| Rosaceae | 11 | 4.5 |
| Apiaceae | 10 | 4.1 |
| Caryophyllaceae | 7 | 2.8 |
| Lamiaceae | Lamiaceae | 2.4 |
| Liliaceae | 6 | 2.4 |
| Salicaceae | 6 | 2.4 |
| Others (60 families) | 93 | 37.2 |
| Total | 248 | 100.0 |
Life-forms
Raunkiaer dormancy forms of the 248 taxa in the survey area were analyzed. The dormancy form with the highest richness was therophytes (Th) in 73 taxa (29.4%), followed by hemicryptophytes (H) in 64 taxa (25.8%), geophytes (G) in 35 taxa (14.1%), and helop hytes (HH) and nanophanerophytes (N) in 17 taxa (6.9%; Table 3). Montane wetlands tend to show a high abundance of hemicryptophytes (Kim et al ., 2011; Ko et al ., 2014), while riverine wetlands such as Yongyangbo Wetlands are at low altitudes and surrounded by farmland, roads, and other facilities, resulting in the appearance of a large number of therophytes, including naturalized plants. The disturbance index at Yongyangbo Wetlands was 33.8%, indicating a more stable habitat than orchards (47.0%, Kim et al .., 2019) or fields (55.0%, Kim et al ., 2015) known to undergo continual human management. It is also a more stable habitat than Hak Reservoir, which is a lacustrine wetland in the DMZ (38.2%, Shin et al .., 2015).
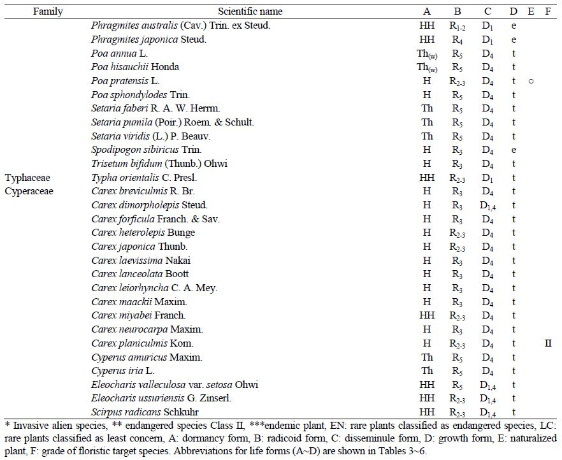
Table 3.
| Dormancy form * | Perennial | Annual | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Dormancy form * | Ch | E | G | H | HH | M | MM | N | HH(Th,Thw) | Th | Th(w) |
| No. of taxa | 11 | 1 | 35 | 64 | 17 | 16 | 14 | 17 | 6 | 32 | 35 |
| Ratio (%) | 4.4 | 0.4 | 14.1 | 25.8 | 6.9 | 6.5 | 5.6 | 6.9 | 2.4 | 12.9 | 14.1 |
*Ch, chamaephyte; G, geophyte; H, hemicryptophyte; HH, hydatophyte; M, microphanerophyte; MM, megaphanerophyte; N, nanophanerophyte; Th, therophyte (summer annual); Th(w): therophyte (winter annual).
The most commonly observed radicoid form was R5(147 taxa, 59.3%) (a non clonal form, such as Erigeron annuus (L.) Pers., Setaria pumila (Poir.) Roem. & Schult., or Inula britannica var. japonica (Thunb.) Franch. & Sav), followed by R3 (49 taxa, 19.7%) that showed short branching rhizomes and clonal growth over a narrow range, such as Eragrostis ferruginea (Thunb.) P. Beauv. or Aster incises Fisch . (Table 4). Among disseminule forms, clitochores (D4, 129 taxa, 52.0%) was the most common, followed by anemochore or hydrochore (D1, 44 taxa, 17.8%) (for which seeds are carried on wind or wa ter) and zoochore (D3, 24 taxa, 9.7%) (for which seeds are carried by sticking to animals or humans) (Table 5). Among growth forms as a way to categorize the state of growth and morphology of aerial parts, the erect form (73 taxa, 29.4%) was the most common, followed by the tussock form (t, 40 taxa, 16.1%) and the pseudo rosette form (ps, 27 taxa, 10.9%; Table 6). Thus, life forms of plants in Yongyangbo Wetlands could be Th R(5)-D4-e. This is a life form composition similar to that of plants in orchards and fields, but different from the life form composition of H-R5-D4-e generally observed for forest plants (Kang et al .,2006).
Table 4.
| Radicoid form* | R1-2 | R2-3 | R2-3(b) | R3 | R3(b) | R3(o) | R3(s) | R3(v) | R4 | R5 | R5(b) | R5(o) | R5(s) | R(s) | R(v) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of taxa | 1 | 29 | 1 | 38 | 2 | 1 | 2 | 6 | 18 | 141 | 1 | 1 | 4 | 2 | 1 |
| Ratio (%) | 0.4 | 11.7 | 0.4 | 15.3 | 0.8 | 0.4 | 0.8 | 2.4 | 7.3 | 56.9 | 0.4 | 0.4 | 1.6 | 0.8 | 0.4 |
*R1 : : widest extent of rhizomatous growth; R2: moderate extent of rhizomatous growth; R3: narrowest extent of rhizomatous growth; R4:clonal growth by stolons and struck roots; R1-2: plants with rhizomatous mutation of R1 and R2;R2-3: plants with rhizomatous mutation of R2 and R3, R(b): Bulb, R(c): Corm, R(o), oblique type, R(s): succulent type, R(v): vertical
Table 5.
| Disseminule form* | D1 | D1,4 | D2 | D2,4 | D3 | D3,2 | D4 | D4,1 | D4,2 | D5,4 |
|---|---|---|---|---|---|---|---|---|---|---|
| No. of taxa | 44 | 9 | 24 | 10 | 24 | 1 | 129 | 4 | 1 | 2 |
| Ratio (%) | 17.8 | 3.6 | 9.7 | 4.0 | 9.7 | 0.4 | 52.0 | 1.6 | 0.4 | 0.8 |
*D1 : widely disseminated by wind or water; D2 : disseminated attachment with or eaten by animals and humans; D3 : disseminated by mechanical propulsion of dehiscence of fruits; D4 : no special modification for dissemination; D5 : not producing seeds; D1,4 : plants with D1 and D4, D2,4 : plants with D2 and D4, D3,2 : plants with D3 and D2, D4,1 : plants with D4 and D1, D4,2 : plants with D4 and D2, D5,4 : plants with D5 and D4
Table 6.
| Growth form* | b | b,e | b-l | b-p | b-ps | e | e,b | l | l-b | p | p-b | p-e | p-l | p-ps | pr | ps | ps-b | r | t | t-p |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of taxa | 16 | 1 | 6 | 4 | 3 | 73 | 12 | 21 | 3 | 2 | 1 | 1 | 1 | 4 | 15 | 27 | 2 | 13 | 40 | 3 |
| Ratio (%) | 6.5 | 0.4 | 2.4 | 1.6 | 1.2 | 29.4 | 4.8 | 8.5 | 1.2 | 0.8 | 0.4 | 0.4 | 0.4 | 1.6 | 6.1 | 10.9 | 0.8 | 5.3 | 16.1 | 1.2 |
*b: branched form; e: erect form; l: liana form; p: procumbent form; pr: partial rosette form; ps: pseudo rosette form; r: rosette form; t: tussock form, b,e: b form and/or e form, b-l: b form with liana stem, b-p: b form with procumbent stem, b-ps: b form with pseudo rosette, e,b: e form and/or b form, l-b: l form with branched form, p-b: p form with branched form, p-e: p form with e form, p-l: p form with liana form, p-ps: p form with pseudo rosette, ps-b: ps form with branched form, t-p: t form with procumbent stem.
Rare and endemic plants
Rare plants are native plants that require special protection because of their decreasing population sizes or habitats. Three taxa of rare plants were observed in Yongyangbo Wetlands: Silene capitata Kom., Polygonatum stenophyllum Maxim. (both of which are classified as endangered species), and Aristolochia contorta Bunge (classified as a species of least concern) (Fig. 2). Of these, S. capitata has been designated by the Ministry of Environment as a Class II endangered wildlife. Forty individuals were observed on roadside cliffs at three points. A relatively large distribution of A. contorta was observed near the farmland and edges of the wetlands.
Fig. 2.

Because Yongyangbo Wetlands are located in the CCZ, it is not a region that can be freely accessed by civilians. However, it can be accessed by tourists who submit an application. The endangered plant S. capitata is located on rocks behind an exploration z one made of wooden decking. Therefore, there is a risk of destruction due to the approach of tourists. To protect S. capitata, it is necessary to take active measures to prevent its destruction, such as installing signposts or a fence to restrict access.
Endemic plants are native plants that are only found in South Korea. Salix koriyanagi Kimura ex Goerz was the only endemic plant taxon observed in the survey area. This species was observed on flat sands of the river terrace where waterways meet upstream of Yongyangbo Wetlands. Salix purpurea var. smithiana Trautv. and S. koriyanagi were co dominant. They formed a unique vegetative landscape that characterized Yongyangbo Wetlands.
Floristic target species and naturalized plants
A total of 17 taxa of florist ic target species were identified in Yongyangbo Wetlands. There was one Class V taxon ( S. capitata ), one Class IV taxon P. stenophyllum ), three Class III taxa (Sinomenium acutum (Thunb.) Rehder & E. H. Wilson, Thalictrum rochebrunianum Franch. & Sav., S. purpurea var. smit hiana), five Class II taxa ( Alisma orientale (Sam.) Juz., Alisma canaliculatum A. Braun & C. D. Bouché, Carex planiculmis Kom., Weigela florida (Bunge) A. DC., Mukdenia rossii (Oliv.) Koidz.), and seven Class I taxa ( Adoxa moschatellina L., A. contorta, Impatiens nolitangere L., Deutzia uniflora Shirai, Salix chaenomeloides Kimura, Hemiptelea davidii>(Hance) Planch., Ulmus davidiana var. japonica (Rehder) Nakai ; Table 7).
Table 7.
| Grade | Family | Scientific name |
|---|---|---|
| Ⅴ | Caryophyllaceae | Silene capitata Kom. |
| Ⅳ | Liliaceae | Polygonatum stenophyllum Maxim. |
| Ⅲ | Menispermaceae | Sinomenium acutum (Thunb.) Rehder & E. H. Wilson |
| Ranunculaceae | Thalictrum rochebrunianum Franch. & Sav. | |
| Salicaceae | Salix purpurea var. smithiana Trautv. | |
| Ⅱ | Alismataceae | Alisma orientale (Sam.) Juz. |
| Alisma canaliculatum A. Braun & C. D. Bouché | ||
| Cyperaceae | Carex planiculmis Kom. | |
| Diervillaceae | Weigela florida (Bunge) A. DC. | |
| Saxifragaceae | Mukdenia rossii (Oliv.) Koidz. | |
| Ⅰ | Adoxaceae | Adoxa moschatellina L. |
| Aristolochiaceae | Aristolochia contorta Bunge | |
| Balsaminaceae | Impatiens nolitangere L. | |
| Hydrangeaceae | Deutzia uniflora Shirai | |
| Salicaceae | Salix chaenomeloides Kimura | |
| Ulmaceae | Hemiptelea davidii (Hance) Planch. | |
| Ulmus davidiana var. japonica (Rehder) Nakai |
There were 26 taxa of naturalized plants observed in the survey area, including Fallopia dumetorum (L.) Holub, Rumex crispus L., Chenopodium album var. spicatum W. D. J. Koch, Lepidium virginicum L., Thlaspi arvense L., and Potentilla supina L. The naturalization index was 10.5%. Annual herbaceous plants (16 taxa) constituted the largest proportion of these, followed by perennial herbaceous plants (eight taxa) and woody plants (two taxa Table 8). Most naturalized plants were observed near roads, where there were frequent human interferences. Three invasive alien species were observed: Ambrosia trifida L., Ambrosia artemisiifolia L., and Humulus japonicus Siebold & Zucc. Although only a s mall number of scattered individuals were observed owing to regular weeding, given that they must have entered from adjacent areas, changes in population sizes for these species need to be monitored continuously.
Table 8.
| Family | Scientific name | Dormancy form |
|---|---|---|
| Asteraceae | Ambrosia trifida L.* | Th |
| Ambrosia artemisiifolia L.* | Th | |
| Carduus crispus L. | Thw | |
| Conyza canadensis (L.) Cronquist | Thw | |
| Erigeron annuus (L.) Pers. | Thw | |
| Galinsoga quadriradiata Ruiz & Pav. | Th | |
| Taraxacum officinale F. H. Wigg. | H | |
| Brassicaceae | Lepidium virginicum L. | Thw |
| Thlaspi arvense L. | Thw | |
| Chenopodiaceae | Chenopodium album var. spicatum W. D. J. Koch | Th |
| Convolvulaceae | Ipomoea purpurea (L.) Roth | Th |
| Euphorbiaceae | Euphorbia maculata L. | Th |
| Fabaceae | Amorpha fruticosa L. | N |
| Melilotus suaveolens Ledeb. | Thw | |
| Robinia pseudoacacia L. | MM | |
| Trifolium pratense L. | H | |
| Trifolium repens L. | Ch | |
| Iridaceae | Iris pseudacorus L. | G |
| Onagraceae | Oenothera biennis L. | Thw |
| Poaceae | Bromus tectorum L. | Th |
| Festuca arundinacea Schreb. | H | |
| Poa pratensis L. | H | |
| Polygonaceae | Fallopia dumetorum (L.) Holub | Th |
| Rumex crispus L. | H | |
| Rosaceae | Potentilla supina L. | Ch |
| Scrophulariaceae | Veronica americana Schwein. ex Benth. | Thw |
*Invasive alien species. Abbreviations for dormancy forms are shown in Table 3.
Conclusion and Suggestions
According to results of this study, a total of 248 taxa were identified in the Yongyangbo Wetlands during the survey, including 72 families and 163 genera. The mo st commonly observed families, in order of descending richness, were Poaceae, Asteraceae, Cyperaceae, and Fabaceae. Their life forms were characterized by Th R5 D4-e, a life form composition similar to that of plants in orchards and fields. Three taxa of r are plants were observed, including S. capitata (a Class II endangered wildlife), 26 taxa of naturalized plants, and 3 taxa of invasive alien species. The disturbance index was 33.8%, indicating that the habitat was more stable than that of orchards or fie lds.
The Yongyangbo Wetlands have been designated as a protected wetland area because of their excellent naturality and integrity obtained through long time preservation with high biodiversity based on various habitat environments. In particular, it is an area where genetic exchange with wildlife populations is possible as it is an ecological linkage channel with the northern area. The activation of eco tourism will introduce economic vitality to the depressed local economy in the DMZ area. In order to maintain its functions and roles, systematic management of these wetlands is necessary because there is concern of threats to habitats of indigenous species and the distribution of alien species caused by disturbances to the environment from military units, farmers, visitors, and river facility management, inter alia.
Acknowledgments
The concept setting and drafting were done by Kwang-Jin Cho. All authors contributed to the methodology, data analysis, manuscript revising and editing. The final version of the manuscript was reviewed and agreed by all authors. Please be advised that all authors included in the thesis do not have any conflict of interest relevant to the academic publication of this research content.
This work was supported by a grant (NIE-A-021-18) from "Basic Survey on Inland Wetlands (’21)" in National Institute of Ecology (NIE) funded by the Ministry of Environment (ME), Republic of Korea.