Introduction
Insects and plants are the basis of ecology and intertwined with each other through complex interactions. The relationship between insects and plant diversity has been discussed in various studies, and positive correlations have been frequently found (Gaston 1992; Procheş et al., 2009; Siemann et al., 1998). Plants provide a variety of benefits to insects, such as food and habitat. Therefore, an integrated investigation of these two taxa is ecologically significant and meaningful. However, although insects and plants are closely related to each other, studies conducted simultaneous surveys of both taxa are relatively insufficient compared to individual surveys.
The ecology of island regions are geographically separated from mainland and often represents specific characteristic patterns of ecosystems, which has attracted the attention of many ecologists (Kim et al., 2016). A general feature of all island systems is that they are separated from the source pool in some distance (Choi & An, 2011; Gillespie & Roderick, 2002). In this context, MacArthur and Wilson (1967) established a famous ecological theory, the theory of island biogeography. According to this theory, the biota on islands is determined by repeated inflow from mainland and extinction on islands, and biodiversity is affected by the size of the island and the distance from mainland, which is the source of species. In general, the closer to mainland and/or the larger size of an island, the greater the number of species would be found on the island (MacArthur & Wilson, 1967). Many follow-up studies on this theory have been conducted, including those that applied the theory to insects or vascular plants in Korea (Jeong & Hong, 2002; Kim et al., 2016; Lee et al., 2008; Park et al., 2013).
In Korea, approximately 4,000 inhabited and uninhabited islands are distributed in the western and southern seas, but compared to the large number of islands, the integrated database and biogeographic study of islands are insufficient (Kim et al., 2016). In this context, insect fauna and vascular plant flora were simultaneously investigated and analyzed for seven islands in Gogunsan archipelago in this study. Each surveyed island was compared through similarity analysis with insects and plants, and the theory of island biogeography was applied. In addition, analyses of insect community indices and naturalized plants of each island were provided.
Materials|Methods
Survey sites
The range of latitude and longitude of Gogunsan archipelago in this study was within 35.794, 126.310 to 35.862, and 126.419. Thirty-year average annual (1981-2010), maximum average annual, minimum average annual temperatures, and precipitation for this reign are 12.8°C, 17.1°C, 9.2°C, and 1,202 mm respectively (Korea Meteorological Administration, 2020). Currently, among the seven islands, Seonyudo, Jangjado, and Daejangdo are connected to mainland by bridges but Gwallido, Bangchukdo, Myeongdo, and Maldo are still separated from mainland.
The surveyed islands are as follows. Seonyu 3 district area has an area of approximately 1.17 km2 in Seonyudo. Jangjado has an area of approximately 0.13 km2, which is connected to the western part of Seonyudo by the Bridge Jangjadaegyo. Daejangdo has an area of approximately 0.337 km2, which is connected to the northern part of Jangjado by the Bridge Daejangdaegyo. Gwallido has an area of approximately 1.652 km2, and is located near the western side of Jangjado and Daejangdo. Bangchukdo has an area of approximately 2.16 km2 the largest island in the surveyed islands. Myeongdo has an area of approximately 0.47 km2, and is located between Bangchukdo and Maldo. Maldo is has an area of approximately 0.36 km2, and is located near the western side of Myeongdo (Fig. 1).
Investigation methods
Surveys were conducted for insects and vascular plants on the seven islands of Gogunsan archipelago from the 19th to the 21st of May, the 14th to the 17th of July, and the 21st, 23rd, and 24th of September 2020 (Table 1). Each of months represents spring, summer, and autumn respectively for survey the three seasons.
The survey hours for each site were approximately 2 hours during each surveyed season for both taxa. For insect surveys, sweeping and brandishing methods were used. Also, for the insects that could identified in the field were counted and recorded in the pictures. The sampled insects were identified using a stereomicroscope (Olympus SZ1145; Olympus, Tokyo, Japan). During the insect identification, various illustrated books and papers (Ahn et al., 2018; Jung & Park, 2014; Lee, 2012; Park et al., 2012) were refered. Entomological Society of Korean and Korean Society of Applied Entomology (1994) and National Institute of Biological Resources (2018) were used for the insect classification system. Vascular plants were photographed or cut and transferred to the laboratory for identification. For the vascular plants identification, Park (2009), Lee (2014a), Lee (2014b), Lee and Lee (2015), Cho et al. (2016), and Kim and Kim (2018) were used. The plant classification system was based on a checklist of vascular plants in Korea (Korea National Arboretum, 2017). The sampled insects and vascular plants were stored as specimens in the Natural History Laboratory of the National Science Museum of Korea.
Analysis methods of insect and vascular plant
For insect community analysis, an equation transformed from the Shannon–Wiener function (H') derived from Margalef's (1958) information theory was used (Pielou, 1966) for the diversity index. McNaughton's equation (1967) was used for the dominance index, Margalef's equation (1958) was used for the richness index, and Pielou's equation (1975) was used for the evenness index. The similarity index was calculated using the equation developed by Sørensen (1948) and analyzed by a multidimensional scaling and clustering methods using IBM SPSS Statistics ver. 21 (IBM Co., Armonk, NY, USA).
Also, to analyze correlations of the distances of each island and the similarities of insect fauna and plant flora on each island, distances among the seven islands were derived from satellite images with the main surveyed areas of each island. To do this, Pearson correlations were calculated using IBM SPSS Statistics.
(ni: number of individuals of the ith species; N: total number of individuals)
(n1: number of dominant species individuals, n2: number of subdominant species individuals, N: total number of individuals)
(S: total number of species; N: total number of individuals)
(H’: species diversity index; N: total number of species)
(S: number of species common to the two sites, Si: number of species in study site i, Sj: number of species in study site j)
To analyze naturalized vascular plants of each surveyed island, the method described by Yim and Jeon (1980) was used to calculate the urbanization indices.
In addition, according to the theory of island biogeography, the correlations among the three kinds of quantitative variables data the numbers of insect species, the numbers of plant taxa, and the areas of island were analyzed using IBM SPSS Statistics with Spearman’s correlation. The three kinds of quantitative variables data are nonparametric data and Spearman’s correlation is generally used instead of parametric Pearson's correlation in two quantitative variables which are not normally distributed (Harris et al., 2008).
The area of each island was determined based on data from the Gunsan Local Culture Encyclopedia website (GLCE, 2020). Seonyudo is originally three islands, and there are Seonyu 1, 2, and 3 districts connected by a sandbar and/or a sand dune. Seonyu 3 district area surveyed in this study was calculated after digitizing the area with QGIS based on satellite images.
Results
The numbers of insect species and individuals, and vascular plant taxa
As a result of this study, a total of 2,817 insect individuals including 264 species were collected (Supplementary Table 1). The total number of vascular plant taxa surveyed was 315 (Supplementary Table 2). Bangchukdo showed the largest numbers of insects species in any surveyed season. Vascular plants taxa also showed the largest number in Bangchukdo among the surveyed islands (Table 2).
Dominant and subdominant insect species by the sites and seasons
For the total individuals of insects surveyed in this study, the dominant species was Spoladea recurvalis, and the subdominant species was Megacopta punctatissima. In September, Spoladea recurvalis was abundant and in May, Megacopta punctatissima was abundant. Jangjado, Daejangdo, and Gwallido have the same dominant speceis Spoladea recurvalis. Also, Myeongdo and Maldo have the same dominant species Nysius plebejus (Table 3).
Community analysis of insect species
As a result of insect community analysis by each island, Bangchukdo had the highest community stability, with the highest diversity, richness and evenness index, and the lowest dominance index (Fig. 2).
The number of naturalized plant and urbanization indices of vascular plant
As a result of the survey of naturalized plants a total of 74 taxa were found. Maldo had the highest urbanization indices among the surveyed sites (Table 4).
Similarity analysis of insect and vascular plant
The similarity index is regarded as a heterogeneous group when the group is less than 20% similar and a homogenous group when the group is 80% or more similar (Ahn & Park, 2012; Whittaker, 1956). Regarding the similarity analysis of insects in this study, all cases were more than 0.2, except for Daejangdo and Maldo (Fig. 3). Bangchukdo and Myeongdo had the highest similarity (0.667). The islands can be divided into two groups based on similarity indices results (Seonyudo, Jangjado, and Daejangdo group and the other islands group).
Regarding the similarity analysis of vascular plants in this study, all cases were more than 0.2 similar (Fig. 4). Bangchukdo and Maldo had the highest similarity (0.474). The islands can be divided into three groups based on similarity indices results (Jangjado, Daejangdo and Gwallido group, Bangchukdo, Myeongdo, and Maldo group and Seonyudo group).
To clarify correlations of the distances among the islands and the similarity indices of the two taxa, we used Pearson's correlation analysis. As a result of analysis, the correlation coefficient between the insect similarities and the inter-island distance was –0.484, confirming the negative correlation (P-value 0.026). Also, in the case of the plant similarities and the inter-island distance, the correlation coefficient was –0.5 (P-value 0.021).
Correlation of the numbers of insect species, vascular plant taxa, and the areas of each island
In this study, Spearman’s correlation was used to apply the theory of island biogeography. For the areas of each island and the numbers of insect species, the correlation coefficient was 0.857 and the P-value (0.014) was less than significant value 0.05. In the case of the numbers of plant taxa, correlation with the areas of the island was not found. Also, the distance between the islands and mainland were not related in both the numbers of insect species and vascular plant taxa. In addition, the numbers of insect species and vascular plant taxa were analyzed to consider the relationship between insect and plant richness. The result showed the correlation coefficient of 0.714 and the P-value (0.071) was larger than the significant value 0.05 but smaller than 0.1 (Table 5).
Discussion
The total number of surveyed insect species and individuals in this study (264 species, 2,817 individuals) was higher than that of uninhabited islands in Sinan-gun, with 122 species and 921 individuals (Cho et al., 2011). These results are assumed to be because the 28 uninhabited islands in Sinan-gun are mainly smaller and the distance from mainland was larger than the islands surveyed in Gogunsan archipelago. The average area of the 28 surveyed uninhabited islands in Sinan-gun is 0.048 km2 which is much smaller than the average area of the seven surveyed islands in Gogunsan archipelago (0.897 km2) (Cho et al., 2011).
Among the insects surveyed in this study, Spoladea recurvalis was the dominant species, with 341 individuals, and the subdominant species was Megacopta punctatissima with 147 individuals. These species were followed by Nysius plebejus, with 127 individuals. A survey of insects in Hallyeo Haesang National Park also confirmed that Nysius plebejus and Megacopta punctatissima were the dominant and subdominant species (Cheong et al., 2005). Also, in comparison with the results of insect surveys on uninhabited islands in Sinan-gun, the subdominant species was Spoladea recurvalis in Lepidoptera (Cho et al., 2011). The host of Nysius plebejus is a very wide range including Asteraceae, and the main host plant of Megacopta punctatissima is Pueraria lobata which can be commonly seen in Korea, and Spoladea recurvalis is a major pest with a very wide host range, so these results seems to be very general (Ahn et al., 2018; Hsu & Srinivasan, 2012; Kim et al., 1994).
Recently, Seonyudo, Jangjado, and Daejangdo in Gogunsan archipelago were connected to mainland by bridges. Jang and Kim (2014) conducted a vascular plants survey in Gogunsan archipelago including the same island (Jangjado) with this study before the constructions of bridges were completed and the result was that fewer vascular plants taxa were surveyed (74 taxa in Jangjado) than in this study (92 taxa in Jangjado). Also, Urbanized index for Jangjado (1.43) were lower than this study (6.23). It is assumed that the invasion of external species and moderate disturbance after the construction of the briges caused an increase in the number of vascular plants species including naturalized plants.
By hierarchical cluster analysis, the Jangjado, Daejangdo group and the Bangchukdo, Myeongdo, and Maldo group were classified by both insects and plants suggesting high correlation between the nearness of distance and similarity indices. Also, multidimention analysis of similiarity indices provided strong relation of nearness of each island intuitively because it shows similar position of islands with actual position in a satellite map in both insect and plant. The high similarity index of insects for Gwallido and Maldo may suggesting the similar environment of two islands.
The high correlation between the numbers of insect species and the areas of each surveyed island was confirmed by Spearman’s correlation coefficient. In the case of plant, the correlation between the areas and the numbers of species was not related in this study, and it is assumed that other factors such as edaphic condition, topography, sampling method, and human interference can be a greater effect in this case. Also, the distance between the island and mainland, and the numbers of species were not related in both taxa. A similar result was found in another study on vascular plant and island biogeography in Southern Sea of Jeollanamdo, Korea, reporting that this may be due to the distances between islands being too close to apply the theory of island biogeography (Kim et al., 2016) which suggesting that more distances between islands should be needed to apply the island biogeography theory. For the case of the study on insect fauna in the west coast of Incheon which includes more remote islands, surveyed islands far from mainland showed less number of insect species than nearer islands (Park et al., 2013).
In this study, we compared the seven islands in Gogunsan archipelago with insect fauna and vascular plant flora. Also, the correlation of the numbers of insect species, vascular plant taxa, and the areas of each island was analyzed. In the future research, it is suggested that considering more factors quantitatively such as a method of sampling, an impact of human interference, and habitat diversity. The results of this study can be used as data related to island ecology and conservation.
References
GLCE (Gunsan Local Culture Encyclopedia) (2020, Retrieved December 11, 2020) Geology and Topography from http://gunsan.grandculture.net/gunsan/toc?search=A1/2
National Institute of Biological Resources (NIBR) (2018, Retrieved December 11, 2020) National Species List of Korea from http://kbr.go.kr
Figures and Tables
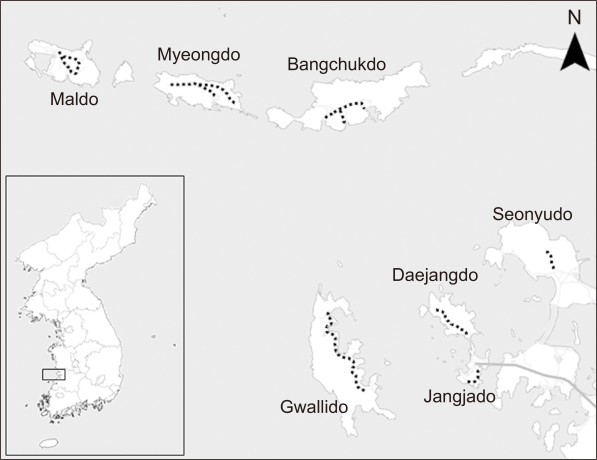
Fig. 2
Community structure analysis of insect by the seven islands in Gogunsan archipelago, 2020. (A) Diversity index. (B) Dominance index. (C) Richness index. (D) Evenness index.
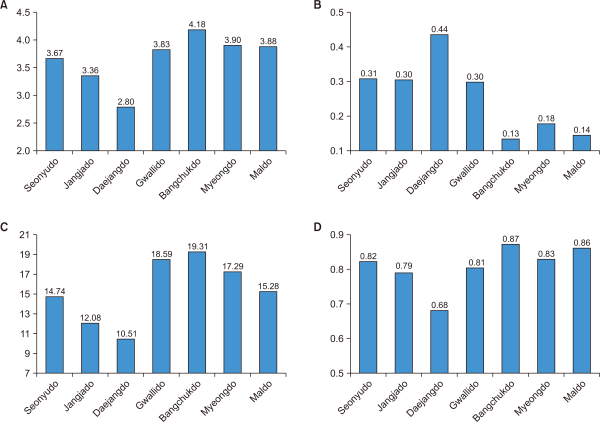
Fig. 3
Comparison of the similarity indices to insect among the sites. UPGMA, Unweighted Pair Group Method with Arithmetic Mean.
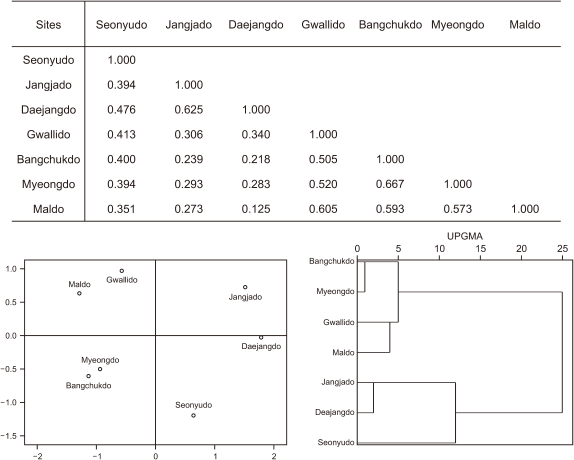
Fig. 4
Comparison of the similarity indices to vascular plant among sites. UPGMA, Unweighted Pair Group Method with Arithmetic Mean.
Table 1
Survey dates on each island in Gogunsan archipelago, 2020
| Season | Site | ||||||
|---|---|---|---|---|---|---|---|
|
|
|||||||
| Seonyudo | Jangjado | Daejangdo | Gwallido | Bangchukdo | Myeongdo | Maldo | |
| Spring | 2020-05-19, 20 | 2020-05-20 | 2020-05-19, 20 | 2020-05-21 | 2020-05-21 | 2020-05-21 | 2020-05-21 |
| Summer | 2020-07-17 | 2020-07-14 | 2020-07-17 | 2020-07-16 | 2020-07-16 | 2020-07-15 | 2020-07-15 |
| Autumn | 2020-09-21 | 2020-09-21 | 2020-09-21 | 2020-09-23 | 2020-09-24 | 2020-09-24 | 2020-09-23 |
Table 2
The results of survey of insect species and plant taxa in Gogunsan archipelago, 2020
| Taxa | Site | ||||||||
|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||
| Seonyudo | Jangjado | Daejangdo | Gwallido | Bangchukdo | Myeongdo | Maldo | Total | ||
| Insect | May | 15 (85) | 27 (77) | 20 (99) | 37 (74) | 31 (107) | 27 (167) | 23 (54) | 92 (663) |
| July | 58 (163) | 28 (93) | 27 (52) | 58 (161) | 62 (188) | 58 (169) | 37 (101) | 142 (927) | |
| September | 32 (72) | 40 (132) | 28 (150) | 50 (251) | 72 (205) | 68 (211) | 56 (206) | 154 (1,227) | |
| Total | 86 (320) | 70 (302) | 61 (301) | 116 (486) | 121 (500) | 110 (547) | 91 (361) | 264 (2,817) | |
| Plant | 66 | 92 | 94 | 131 | 164 | 135 | 140 | 315 | |
Table 3
Dominant/subdominant insect species by the seasons and sites in Gogunsan archipelago, 2020
| Season | Site | |||||||
|---|---|---|---|---|---|---|---|---|
|
|
||||||||
| Seonyudo (Domiant/ subdominant species) |
Jangjado (Domiant/subdominant species) |
Daejangdo (Domiant/subdominant species) |
Gwallido (Domiant/subdominant species) |
Bangchukdo (Domiant/subdominant species) |
Myeongdo (Domiant/subdominant species) |
Maldo (Domiant/subdominant species) |
Total (Domiant/subdominant species) |
|
| May | Megacopta punctatissima/ Aulacophora nigripennis |
Megacopta punctatissima/ Syrphus ribesii, Coccinella septempunctata |
Melanostoma mellinum/ Megacopta punctatissima |
Megacopta punctatissima/ Aelia fieberi |
Lasius japonicus/ Metasyrphus corollae |
Nysius plebejus/ Pachygrontha antennata |
Megacopta punctatissima/ Coccinella septempunctata |
Megacopta punctatissima/ Coccinella septempunctata |
| July | Nylanderia flavipes/ Pagria signata |
Rivellia basilaris/ Aphaenogaster tipuna |
Aphaenogaster tipuna/ Camponotus vitiosus, Pagria signata |
Chauliops fallax/ Monolepta pallidula |
Basilepta fulvipes/ Cyphononyx fulvognathus |
Bifurcanomala aulax/ Ischnura asiatica |
Nysius plebejus/ Eretes griseus |
Basilepta fulvipes/ Bifurcanomala aulax |
| September | Bothrogonia ferruginea/ Cardipennis sulcithorax |
Spoladea recurvalis/ Oecanthus longicauda |
Spoladea recurvalis/ Aphaenogaster tipuna |
Spoladea recurvalis/ Parnara guttata |
Spoladea recurvalis/ Nysius plebejus |
Spoladea recurvalis/ Bothrogonia ferruginea |
Locusta migratoria migratoria/ Liorhyssus hyalinus |
Spoladea recurvalis/ Nysius plebejus |
| Total | Megacopta punctatissima/ Aulacophora nigripennis, Bothrogonia ferruginea |
Spoladea recurvalis/ Rivellia basilaris |
Spoladea recurvalis/ Melanostoma mellinum |
Spoladea recurvalis/ Nysius plebejus, Parnara guttata |
Basilepta fulvipes/ Spoladea recurvalis |
Nysius plebejus/ Pachygrontha antennata |
Nysius plebejus/ Locusta migratoria migratoria |
Spoladea recurvalis/ Megacopta punctatissima |
Table 4
The number of naturalized plant, and urbanization indices for the surveyed sites in Gogunsan archipelago, 2020
| Division | Site | |||||||
|---|---|---|---|---|---|---|---|---|
|
|
||||||||
| Seonyudo | Jangjado | Daejangdo | Gwallido | Bangchukdo | Myeongdo | Maldo | Total | |
| Naturalized plant | 6 | 20 | 6 | 28 | 28 | 16 | 34 | 74 |
| Urbanization index | 1.87 | 6.23 | 1.87 | 8.72 | 8.72 | 4.98 | 10.59 | 23.05 |