Introduction
Amphibians are considered to have been the first tetrapods to venture onto land and have been challenged by and adapted to extreme environments since ancient times (Shubin & Jenkins, 1995; Wake & Koo, 2018; Wells, 2010). The order Anura (frogs and toads) accounts for approximately 88% of all amphibian species and is a particularly diverse vertebrate group (AmphibiaWeb, 2021; Wake & Koo, 2018). Most anuran species emerge as tadpoles from eggs laid in water before undergoing metamorphosis and coming onto land in their adult form. Adult anurans have diverse lifestyles and occupy a wide range of terrestrial and freshwater environments. They occur in permanent ponds, marshes, streams, and lakes and can be found in rainforests, deserts, alpine regions, and even Alaskan permafrost (Wells, 2010). Some species are mostly terrestrial, while others live almost all their lives in an aquatic environment, and some semi-aquatic species alternately use a variety of terrestrial and aquatic environments (Moen & Wiens, 2017). In different environments, they survive with morphologies independently derived across several clades (Hall, 2008; Wells, 2010).
Unlike the two other amphibian groups, Caudata and Gymnophiona, the anurans lost their tails during evolution and now rely on their forelimbs and hindlimbs for locomotion (Emerson, 1978; 2013; Wells, 2010). Some species jump when they are on land using their lower body, including their hind legs, whereas others swim in an aquatic environment almost all their lives. Other anurans use their forelimbs and hindlimbs to walk, with only a few jumps, and can even climb trees using round adhesive discs on their fingers and toes (Enriquez-Urzelai et al., 2015; Mendoza et al., 2020; Moen & Wiens, 2017). There are also species with unique locomotor patterns, such as flying frogs (Emerson & Koehl, 1990). Some species burrow with their hindlimbs to survive in dry conditions (Moen et al., 2013; Vidal‐García & Keogh, 2015). Ecological factors, such as microhabitats, predator interactions, and defense mechanisms, can alter anuran behavioral traits, e.g., locomotor performance (Buttimer et al., 2020; Mendoza et al., 2020; Vega-Trejo et al., 2014). To understand the trade-off between behavioral adaptation and morphological function, it is necessary to quantify the morphological differences according to the ecological traits of various anuran species.
Morphological differences related to various ecological traits of anurans can affect their locomotor performance and mode. Commonly, locomotor performance is related to body size or body weight in frogs (Emerson, 1978; Nauwelaerts et al., 2007). Additionally, these locomotory performances are closely related to the anatomical shapes as well as the external traits. In the lower body skeleton in frogs, a narrow pelvis, long ilium and urostyle, and short presacral vertebrae result in improved jumping performance (Emerson, 1978; Nauwelaerts et al., 2007; Zug, 1972). The morphology of the forelimbs or hindlimbs is also related to the locomotion mode of frogs (Enriquez-Urzelai et al., 2015). The length and ratio of the hindlimb, including the femur and tibiofibular, can change the propulsion and take-off speed for jumping. In fact, this ratio differs according to the various locomotion modes, such as arboreal jumpers, jumpers, swimmers, walkers, hoppers, and burrowers (Enriquez-Urzelai et al., 2015). The forelimbs do not have great direct propulsion but are used to soften the impact on landing (Nauwelaerts & Aerts, 2006). Indirectly, short forelimbs can move the center of mass in frogs backward and increase acceleration when jumping (Emerson, 2013).
In this study, we attempted to quantify the differences in the skeletal shape of four anuran species (Japanese tree frog, Dryophytes japonicus; Japanese wrinkled frog, Glandirana rugosa; black-spotted pond frog, Pelophylax nigromaculatus, and American bullfrog, Lithobates catesbeianus) commonly found in Korea. The four species have ecological differences in body size, locomotor mode by microhabitat, trophic position, and predator evasion tactics. We organized this information through our data and bibliography. The skeletal images of the four species were obtained using radiographic technology, and the skeletal differences of the lower body and limbs were quantified by both landmark-based geometric morphometrics and traditional methods of morphology comparison.
Materials|Methods
Field investigation
Four species of anurans, Japanese tree frogs (D. japonicus), black-spotted pond frogs (P. nigromaculatus), Japanese wrinkled frogs (G. rugosa), and American bullfrogs (L. catesbeianus), were collected by hand from May 2019 to July 2019 in Gongju City. We captured ten frogs from four different sites to exclude habitat differences. All frogs were sexually mature, and the male frogs were distinguished by confirmation of the vocal sac and nuptial pad on the first finger, which are secondary sexual characteristics. Among the individuals that did not have male frog characteristics, animals aged 2-3 years or older were judged to be female. Individuals aged 1-2 years were judged to be immature and excluded from the analysis. As a result, a total of 8 frogs from each species were used for morphological analysis.
Measurement of physical conditions
The frogs’ age was analyzed using skeletochronology, which counts the lines of arrested growth (LAGs) in cross-sections of the phalanges. The animals were anesthetized with ice-cold water, which reduces neural activity. Then, the second-right front fingers and the third-right hind toes were collected from each individual. We used 5% nitric acid (HNO3) to decalcify the phalanges and washed the sample with deionized water. Next, a Spin Tissue Processor STP 120 (Myr, Tarragona, Spain) was used to carry out paraffin embedding. The tissue sections were cut to a 12-µm thickness with a Thermo-Shandon Finesse ME Microtome (Thermo Fisher Scientific, Waltham, MA, USA), and Harris’ hematoxylin was used to stain the tissue. The sample was placed on a slide and covered with a glass coverslip. The number of LAGs in each individual was confirmed using a DP72 microscope (Olympus, Tokyo, Japan) at 100× magnification. Experimental procedures on the animals were conducted per the regulations and with the approval of the Experimental Animal Ethics Committee of Kongju National University (KNU_2019-01).
Body size and weight were measured to record the basic physical information of each species, along with age. The snout-vent length (SVL), a measure of amphibian body size, was measured as 0.01 mm units using a digital caliper (IP54 SHAHE Digital Caliper, Zhejiang, China). The body weight was measured as 0.01 g units using a digital balance. The SVL was compared using a one-way analysis of variance (ANOVA), and the body weight and age were compared using the Kruskal–Wallis test to confirm the significant differences in basic physical condition among the four species. GraphPad Prism version 8.0, for Windows (GraphPad Software, San Diego, CA, USA) was used for statistical analysis, and all statistical differences were considered significant at P<0.05.
Ecological traits of the four anuran species
We summarized the ecological traits and locomotor modes of the four species based on existing literature and our data. The Japanese tree frog, belonging to the Hylidae family, is a terrestrial amphibian and an arboreal species with round adhesive discs on the tips of its fingers and toes (AmphibiaWeb, 2017). Its locomotory mode is typical of arboreal jumpers and enables it to climb trees and grasses (Enriquez-Urzelai et al., 2015; Jorgensen & Reilly, 2013). Japanese tree frogs have the lowest trophic position among the four species (Park & Do, 2019) and are exposed to predators such as bullfrogs in their habitat (Park & Do, 2020). For defense against predators, they use camouflage, changing the dorsal color to match the background (Kang et al., 2016).
The Japanese wrinkled frog, belonging to the Ranidae family, is a semi-aquatic amphibian (AmphibiaWeb, 2019a). Although previous studies have classified these species as terrestrial jumpers (Enriquez-Urzelai et al., 2015; Jorgensen & Reilly, 2013), most semi-aquatic frogs alternate between aquatic and terrestrial environments and perform both swimming and jumping with the same morphological traits (Nauwelaerts et al., 2007). Therefore, we described them as jumpers and swimmers in the present study. The trophic position of the Japanese wrinkled frog is in the middle of the four frog species (Park & Do, 2019). As tadpoles and adults, they produce odorous skin secretions that effectively defend them against predation (Takahara et al., 2011; Yoshimura & Kasuya, 2013).
Black-spotted pond frogs have several ecological traits similar to those of the Japanese wrinkled frogs. This frog belongs to the Ranidae family and is a semi-aquatic amphibian (AmphibiaWeb, 2007). Like the Japanese wrinkled frog, the locomotor mode corresponds to the jumper and swimmer type (Enriquez-Urzelai et al., 2015; Jorgensen & Reilly, 2013). In addition, they have a trophic level similar to the Japanese wrinkled frog (Park & Do, 2019). However, unlike the Japanese wrinkled frog, they have no known defense against predation other than escape. Therefore, they evade predators through their strong jumping or swimming abilities.
The American bullfrog has the largest body size among the four anuran species. Like the Japanese wrinkled frog and black-spotted pond frog, it belongs to the Ranidae family, and its locomotor modes are the jumper and swimmer type (AmphibiaWeb, 2019b; Enriquez-Urzelai et al., 2015; Jorgensen & Reilly, 2013). This frog is an apex predator with the highest trophic position among the four species (Park & Do, 2019).
In addition to the body size differences we measured directly, the differences in skeletal shape were analyzed with reference to the ecological traits summarized here: locomotor mode (arboreal jumper, jumper, and swimmer), microhabitat (terrestrial and semi-aquatic frog), and defense mechanisms (camouflage color, skin secretion, jumping and swimming, and apex predator).
Skeletal imaging
The frogs were euthanized by pithing after the body size and weight had been measured and stored as fixed samples in 70% ethanol. The samples were prepared in the same posture for morphological analysis. Styrofoam was fixed to the bottom of a plastic container with 70% ethanol, and the frogs were placed on it. A pin was then used to fix the frog posture. After 3 days, the frog posture was completely fixed, and the frogs were removed from the Styrofoam plate, labeled, and stored separately.
The skeletal shape of the fixed object was photographed using radiographic techniques. Skeletal images were taken using dual-energy X-ray absorptiometry (DEXA; Medikors InAlyzer, Seongnam, Korea) for frogs over 10 g, such as American bullfrogs, black-spotted pond frogs, and female Japanese wrinkled frogs. In the case of frogs weighing less than 10 g, such as Japanese tree frogs and the males of Japanese wrinkled frogs, the image of the skeleton was photographed using a micro-CT instrument (PerkinElmer, Waltham, MA, USA), located at the Korea Basic Science Institute (Gwangju, Korea). The collected skeletal images were used to confirm the difference in the skeletal shape of the lower body and limbs among the four anuran species.
Analysis of skeletal shape
Landmark-based geometric morphometrics was used to compare the skeletal shape of the lower body among the four species of anurans. The skeletal shape was digitized using TpsDig software (Rohlf, 2006). A total of eight landmarks were designated in the skeletal shape of the lower body: (L1) the anterior tip of the urostyle, (L2) the anterior left tip of the ilium, (L3) the anterior right tip of the ilium, (L4) the posterior tip of the urostyle, (L5) the posterior right tip of the pelvis, (L6) the posterior tip of the ischium, (L7) the posterior left tip of the pelvis, (L8) and the anterior tip of the pelvis (Fig. 1).
We used the MorphoJ software (version 1.07a; Faculty of Life Sciences, University of Manchester, Manchester, UK) to convert the digitized landmark coordinates into Procrustes coordinates. Then, canonical variate analysis (CVA) was used to compare the variations in skeletal shape that distinguished the four species of anurans by minimizing the difference within the groups and maximizing differences among the groups. The difference in skeletal shape was visualized through a rectangular grid with the landmark vectors and a wireframe graph of the canonical variate 1 (CV1) and CV2 axes. The morphological differences among the four species were also compared using discriminant function analysis (DFA), which determines the classification function using Fisher’s classification rule. Significant differences among the four species were analyzed by permutation tests with 1000 permutations. Additionally, the Mahalanobis distance by DFA was computed for the four species. We used a graph that matched the average shape of the four species of anurans with the average shape of each species and confirmed the morphological distance among the four species by Mahalanobis distance.
Comparison of the proportions of forelimbs and hindlimbs
If the angle of a limbs is not perfectly fixed, it may not be suitable for use in landmark analysis. Therefore, the shape of the limbs was analyzed by measuring the length of each skeleton and calculating the proportions. First, the length of the femur, tibiofibular, humerus, and radio-ulnar were measured using ImageJ software version 1.8.0 (Schneider et al., 2012). Then, the tibiofibular to femur ratio (T/F ratio) was computed to confirm the differences among species with different locomotor modes. Additionally, the forelimb (radio-ulnar and humerus) to hindlimb (tibiofibular and femur) ratio (FL/HL ratio) was calculated to identify the difference between terrestrial frogs and semi-aquatic frogs. One-way ANOVA was used to compare the differences in T/F and FL/HL ratios among the four species of anurans using GraphPad Prism version 8.0 for Windows. All statistical differences were considered significant at P<0.05.
Results
Physical condition of the four species
The SVL (F=181) and body weight (W=29.09) were significantly different (P<0.05) among the four species of anurans (Table 1). In particular, SVL was significantly different (Tukey’s post hoc test, P<0.05) among all four species. The Japanese tree frog (D. japonicus) had the lowest SVL (Tukey’s post hoc, P<0.05), whereas the SVL of the American bullfrog (L. catesbeianus) was the highest (Tukey’s post hoc, P<0.05). The body weight of the Japanese tree frog was not different (Dunn’s post hoc, P>0.05) from the weight of the Japanese wrinkled frog (G. rugosa) but was lower (Dunn’s post hoc, P<0.05) than that of the black-spotted pond frog (P. nigromaculatus) and American bullfrog. Japanese wrinkled frogs showed no significant difference (Dunn’s post hoc, P>0.05) in body weight from the black-spotted pond frog but had lower (Dunn’s post hoc, P<0.05) body weight than that of the American bullfrog. The body weight of black-spotted pond frogs was not different (Dunn’s post hoc, P>0.05) from the weight of American bullfrogs. The ages of the four species did not differ (W=6.323, P>0.05) in this study.
Differences in the skeletal shape
The CVA showed two major morphological variations based on CV1 and CV2 among the four species of anurans (Fig. 2). For example, CV1 explained 75.94% of skeletal shape variance, and an individual with a higher CV1 had a narrower pelvis and slightly shortened urostyle. On the contrary, CV2 explained 16.13% of skeletal shape variance, and an individual with a higher CV2 had a wider pelvis, longer ilium, and much shorter urostyle. Japanese tree frogs and black-spotted pond frogs had the highest CV1 values, whereas the American bullfrogs and Japanese wrinkled frogs had the lowest CV1 values. The CV2 values of Japanese tree frogs and Japanese wrinkled frogs were the highest, whereas the American bullfrogs and black-spotted pond frogs had the lowest CV2 values. Procrustes distance and Mahalanobis distance from 1,000 permutation rounds in CVA explained that the skeletal shapes of the four species were significantly different (P<0.05) among all groups. Similarly, DFA showed that the morphological differences among the four species of anurans were independent. After the 1000-permutation test using T-square, all individuals were correctly reclassified (P<0.05) by the discriminant functions (Table 2).
Morphological distance of the skeletal shape
The morphological distance was significant (Mahalanobis distance by DFA, P<0.05) among the four species. Black-spotted pond frogs and American bullfrogs were morphologically the closest. The Japanese tree frogs were also morphologically close to these two species, and the Japanese wrinkled frogs had the most morphologically distance.
Among the four species, black-spotted pond frogs showed a narrow pelvis, average ilium, and long urostyle. American bullfrogs had a wider pelvis, short ilium, and the longest urostyle, whereas Japanese tree frogs had a small pelvis, long ilium, and short urostyle. Japanese wrinkled frogs had a relatively wider pelvis, average ilium, and a short urostyle (Fig. 3).
Comparison of the limb proportions
The T/F ratio varied (F=23.3, P<0.001) among the anurans (Fig. 4A). The T/F ratio of the Japanese tree frog, an arboreal jumper and terrestrial amphibian, was the lowest (Tukey’s post hoc, P<0.05) of the four species. In contrast, the T/F ratio did not differ significantly (Tukey’s post hoc, P>0.05) among the other three species, which are semi-aquatic and jumper/swimmer frogs (Japanese wrinkled frogs, black-spotted pond frogs, and American bullfrogs).
Similarly, the FL/HL ratio also differed (F=6.797, P<0.001) among the species (Fig. 4B). Japanese tree frogs had the highest (Tukey’s post hoc, P<0.05) FL/HL ratio of the four. However, among the other three species of frogs, the FL/HL ratio was not significantly different (Tukey’s post hoc, P>0.05).
Discussion
In this study, we compared the skeletal shape of four species of anurans according to their locomotor mode and ecological traits. The skeletal shapes of the lower body of the four species of anurans were all independently and completely isolated. The analysis showed that the Japanese wrinkled frog was the most morphologically distinct, and the black-spotted pond frog and American bullfrog had the most similar morphologies. The ratio of the limbs for the Japanese tree frogs was considerably different from that of the other three species. We believe that the differences in the skeletal shape of the four species relate to the ecological traits of each species.
The shapes of the pelvis, ilium, and urostyle were different for each species. Although these shapes do not unconditionally determine locomotor performance, they nevertheless affect it (Emerson, 1978; Nauwelaerts et al., 2007; Zug, 1972). In our results, the skeletal shape of the lower body seems to differ according to body weight and predator defense mechanisms. The black-spotted pond frog could achieve the best locomotor performance of the four species, followed by the American bullfrog. The Japanese tree frog and Japanese wrinkled frog had traits representing low locomotor performance. The skeletal shape of the lower body and relatively high locomotor performance of black-spotted pond frogs and American bullfrogs is thought to be due to their higher body mass compared to the other two species. Body mass is a critical variable in explaining variations in the jumping power of anurans, and high body mass can reduce jumping power per body mass (Mendoza et al., 2020). Therefore, it seems that species with a greater weight may need a skeletal shape with better locomotor performance. Additionally, the black-spotted pond frogs exhibited the trait with the highest locomotor performance, possibly because they have no predator defense mechanism other than jumping and swimming. The American bullfrog is a predator with a high trophic position in South Korea (Park & Do, 2019), with few known predators; therefore, anti-predator strategies relying on locomotor performance may be relatively less useful. Japanese wrinkled frogs use odorous skin secretions (Takahara et al., 2011; Yoshimura & Kasuya, 2013), and Japanese tree frogs use a camouflage pattern (Kang et al., 2016) for defense. On the contrary, although black-spotted pond frogs occupy a trophic position similar to that of Japanese wrinkled frogs (Park & Do, 2019), they seem to have this skeletal shape to evade predators through strong locomotor performance because they have no other defense mechanism.
The morphology of the limbs in anurans is greatly influenced by their microhabitat and locomotor mode (Enriquez-Urzelai et al., 2015; Vidal-García & Keogh, 2017). In our study, the Japanese tree frog, an arboreal jumper and terrestrial frog, the tibiofibular is shorter than the femur, unlike the other three species. In general, hindlimb length can increase locomotor performance (Rebelo & Measey, 2019). In particular, strong jumpers tend to have a T/F ratio greater than 1, and weak jumpers have a ratio close to or less than 1 (Zug, 1972). These hindlimb traits are known to be markedly different among anuran species with different locomotor modes (Enriquez-Urzelai et al., 2015). Likewise, the ratio of forelimbs to hindlimbs showed a similar result. Unlike the other three species, Japanese tree frogs had a high ratio of forelimb lengths. The short forelimb of frogs positions the center of gravity toward the back and enhances jumping performance (Emerson, 2013). This limb morphology shows that Japanese tree frogs may be weaker jumpers or swimmers than the other three species. Unlike semi-aquatic frogs with excellent swimming and jumping abilities, the Japanese tree frog is a species that climbs grass and trees. In addition, because Japanese tree frogs use camouflage colors to hide from predators (Kang et al., 2016), they may be safer from predators if they remain stationary rather than jumping or swimming (Morey, 1990). Therefore, unlike the other three species, Japanese tree frogs do not seem to have to be strong jumpers or swimmers.
Differences in morphology among species can help elucidate the ecological and behavioral traits of organisms. However, it is challenging to quantify and analyze these differences. We used both landmark-based geometric and traditional morphometric methods to compare the skeletal shape of anurans in this study. In the past, the morphology of organisms was analyzed and compared using traditional morphometric methods, such as multivariate morphometric methods based on size data (Marcus, 1990). These methods can capture size information well, but there is less information on shape (Bookstein et al., 1985; Rohlf & Marcus, 1993). To compensate for this, a new shape analysis tool, geometric morphometric analysis, was introduced in the late 1980s (Bookstein, 1989; 1997; Rohlf & Marcus, 1993). This tool provides attractive and intuitive visualization data by graphically representing the results of shape variation. It also allows the comparison of shape changes by statistical quantification (Adams et al., 2004). However, the method is restricted to use within a limited number of taxa or species because it can only analyze parts with the same points. The use of landmarks with the same points within different taxa or species may not be sufficient to describe subtle features (Tatsuta et al., 2018). As such, each of these two methods has advantages and disadvantages. For morphological analysis, setting the analysis method according to the characteristics of each trait or using them to complement each other can help quantify the more detailed trait changes.
References
AmphibiaWeb (2007, Retrieved November 20, 2021) Pelophylax nigromaculatus: Dark-Spotted Frog from https://amphibiaweb.org/species/5109
AmphibiaWeb (2017, Retrieved November 20, 2021) Hyla japonica: Japanese Tree Frog from https://amphibiaweb.org/species/832
AmphibiaWeb (2019a, Retrieved November 20, 2021) Glandirana rugosa: Wrinkled Frog from https://amphibiaweb.org/species/5138
AmphibiaWeb (2019b, Retrieved November 20, 2021) Rana catesbeiana: Bullfrog from https://amphibiaweb.org/species/4999
AmphibiaWeb (2021, Retrieved November 20, 2021) Browse by Family from https://amphibiaweb.org/lists/index.shtml
Figures and Tables
Fig. 1
X-ray bone image of male American bullfrog (Lithobates catesbeianus) obtained by dual energy X-ray absorptiometry. Eight landmark point that represented the skeletal shape of lower body. Forelimbs were consisted of radio-ulna and humerus, and hindlimbs were consisted of tibiofibular and femur.
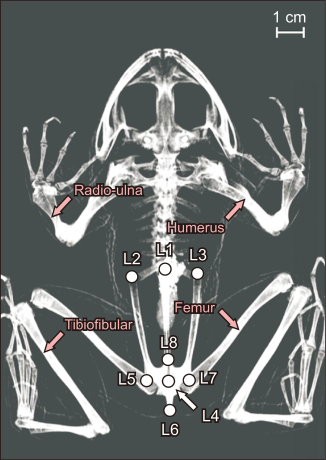
Fig. 2
The variation of skeletal shape among four species anurans from canonical variate analysis (CVA). Scatterplot consisting of canonical variate 1 (CV1) and CV2 shows that four species anurans had different skeletal shape in lower body. Circles in the deformation grids and the grey wireframe graphs indicate the skeletal shape of the individuals with the lowest CV value. Sticks in the deformation grids and the black wireframe graphs indicate the change of skeletal shape by increase of CV value.
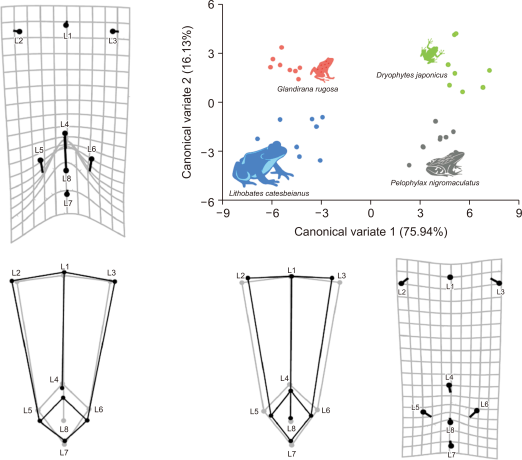
Fig. 3
Mahalanobis distance of skeletal shape among four species anurans obtained by discriminant function analysis (DFA) after the 1000-permutation rounds. The grey wireframe graphs represent mean skeletal shape of four anuran species, and the black wireframe graphs represent mean skeletal shape of each anuran species. All four species showed significant morphological distance (P<0.05).
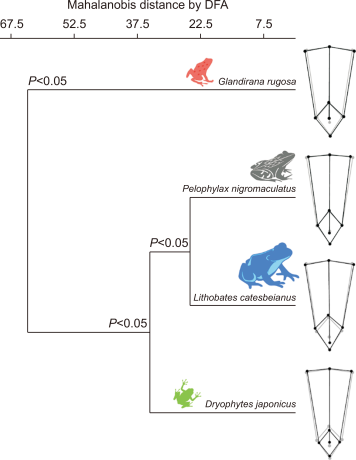
Fig. 4
The comparison in tibiofibular to femur (T/F) ratio (A) and forelimb to hindlimb (FL/HL) ratio (B) among four species anurans. Box plots show the mean (central square dot), median (central band), 25th and 75th percentiles (bottom and top of boxes), range within 1.5 interquartile (IQR, bottom and top of line). Significant differences (P<0.05) were determined using a Tukey’s Post Hoc test from one-way ANOVA test, and are represented with the difference of uppercase letters.
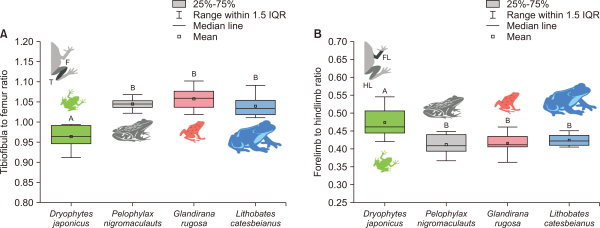
Table 1
The value of snout-vent length (SVL), body weight, and age of four species frogs
| Species | N | SVL (mm)* | Body weight (g)* | Age |
|---|---|---|---|---|
| Dryophytes japonicus | 8 | 33.0±2.2a | 3.4±0.4ab | 3.5±0.5 |
| Glandirana rugosa | 8 | 50.6±8.6b | 10.4±4.4bc | 4.0±0.9 |
| Pelophylax nigromaculauts | 8 | 76.5±9.4c | 32.8±10.8cd | 3.3±0.5 |
| Lithobates catesbeianus | 8 | 150.7±17.6d | 352.4±110.4de | 4.1±0.8 |
Table 2
The discriminant function in the skeletal shape of lower body from four species anurans obtained by discriminant function analysis after the 1000-permutation test using the T-square
| From discriminant function | ||||
|---|---|---|---|---|
|
|
||||
| Dryophytes japonicus | Glandirana rugosa | Pelophylax nigromaculauts | Lithobates catesbeianus | |
| Dryophytes japonicus | 8 (100%) | 0 | 0 | 0 |
| Glandirana rugosa | 0 | 8 (100%) | 0 | 0 |
| Pelophylax nigromaculauts | 0 | 0 | 8 (100%) | 0 |
| Lithobates catesbeianus | 0 | 0 | 0 | 8 (100%) |