Introduction
Lucanidae is one of the basal clade of Scarabaeoidea (Smith et al., 2006), which consist of about 1,400 species from all around world (Fujita, 2010). Among them, Prosopocoilus is one of the biggest genera within Lucanidae. Psalidoremus is one of the subgenus which consists of five species and eleven subspecies through East Asia. Psalidoremus distributes from Taiwan, Japan, Korea and north eastern part of China confirmed by Japanese and Chinese entomologists (Fujita, 2010; Huang & Chen, 2013; Mizunuma & Nagai, 1994). Each species does not cover distribution range of other species, which present strong endemism to their distribution. Most of advance studies focus on the new species or subspecies from this taxon (Adachi, 2009; 2014; Fujita, 2009; Matsuoka & Takatoji, 2010; Shimizu & Murayama, 1998; 2004), therefore, this paper suggests paleogeographical assumption and hypothesis of how they diverged from common ancestors. So far, phylogeographical patterns illustrated by the distribution of each species and divergence were assumed by morphological tendencies especially in male mandibles with assumption from geographical patterns of under sea level.
In these processes, three priori assumptions are critical: 1) Currently recognized taxa are monophyletic; 2) diversification of each subspecies negligibly small compared to its between-island diversification; 3) recognizable speciation occurred by allopatric speciation. These assumptions are adapted and modified from assumptions of Ota which study on the speciation of Amphibians and Reptiles from Ryukyu Archipelago (Ota, 1998).
Materials|Methods
Study design
This study designed by following process. First of all, distributional data of subgenus Psalidormus species has been organized in Table 1.
Data in Table 1 were used in this research with referring global Biodiversity Information Facility (GBIF) and each specimen holding institutions. The distributional data has been referred to occurrence data of the information uploaded to GBIF, and distributional data of key references stag beetle researchers such as Mizunuma, Nagai, Fujita, Huang, and Chen.
After distribution check, morphological data were analyzed by considering male morphology of each species. Especially head structures of male are distinctly differ in each species. Mandibular structure and shape of clypeus is the key morphology to its taxonomy. In mandibular structures, composition of large inner tooth and apical tooth group are considerable. Clypeus shape is organized in three types which considered as gradual changes in speciation process. Female morphology was not analyzed in here because of morphological difference between each species not remarkable as males.
Also, Bathymetric map under sea surface were drawn by author based on open street map to avoid copyright issues. Three bathymetric elevations which respectively 0 m, –750 m, –1,200 m, were used regard species boundary of distribution.
After all, hypothetical speciation history established based on information as mentioned above.
Materials
Materials used in this research mostly stored in the storage of National Science Museum Korea, National Taiwan Museum, and GBIF occurrence data set ( https://doi.org/10.15468/39omei).
Methods
Habitus photograph were taken by Nikon D7100 Digital Single Lens Reflex Camera (Nikon, Tokyo, Japan) with Tamron 60 mm Macro lens which mounted on Wemacro macro rail set with lighting stands. Photo of each specimen were taken 30 to 40 by each stop and merged together in Zerene Stacker program (Zerene Stacker, Richland, WA, USA) to maximize resolution. Maps of each figure were drawn by author by using PaintTool SAI (Systemax, Port Washington, NY, USA). Male morphological structures labeled as Table 2 for the comparison of each trait. Seven morphological traits have been assessed in the IBM SPSS statistics program (ver. 28.0; IBM Co., Armonk, NY, USA) by hierarchical cluster analysis method.
Results
By referring occurrence data of each specimen and dataset, all species of subgenus Psalidoremus could be confirmed as endemic to each distribution. The geographical patterns of the Psalidoremus species has been displayed on Fig. 1. And brief taxonomic account follows.
Taxonomic accounts
Insecta
Scarabaeoidea
Lucanidae Latreille, 1804
Genus Prosopocoilus Westwood, 1845
Subgenus Psalidoremus Motschusky, 1862
Species Prosopocoilus (Psalidoremus) inclinatus (Motschulsky, 1857)
Species Prosopocoilus (Psalidoremus) motschulskii (Waterhouse, 1869)
Species Prosopocoilus (Psalidoremus) dissimilis (Boileau, 1898)
Species Prosopocoilus (Psalidoremus) hachijoensis Nomura, 1960
Species Prosopocoilus (Psalidoremus) pseudodissimilis Kurosawa, 1976
Prosopocoilus (Psalidoremus) inclinatus (Motschusky, 1857) (Fig. 1A)
Body length: male 21-72 mm, female 25-42 mm
Distribution: Korea, Japan, China, Russia
Subspecies: This species consist of seven subspecies. P. inclinatus inclinatus (Motschusky, 1857) P. inclinatus mishimaiouensis Shimizu et Murayama, 1998, P. inclinatus kuchinoerabuensis Shimizu et Murayama, 1998, P. inclinatus kuroshimaensis Shimizu et Murayama, 2004, P. inclinatus miyakejimaensis Adachi, 2009, P. inclinatus mikuraensis Matsuoka et Takatoji, 2010, P. inclinatus yakushimaensis Adachi, 2014.
Prosopocoilus (Psalidoremus) hachijoensis Nomura, 1960 (Fig. 1B)
Body length: male 24-59 mm, female 26-38 mm
Distribution: Japan (Hachijo Island [Is.])
Subspecies: This species does not have subspecies.
Prosopocoilus (Psalidoremus) dissimilis (Boileau, 1898) (Fig. 1C)
Body length: male 26-79 mm, female 25-40 mm
Distribution: Japan (Okinawa Iss., Amami Iss., Tokara Iss.)
Subspecies: This species consist of seven subspecies. P. dissimilis (Boileau, 1898), P. dissimilis elegans (Inahara, 1958), P. dissimilis okinawanus Nomura, 1962, P. dissimilis makinoi Ichikawa et Fujita, 1985, P. dissimilis okinoerabuensis Ichikawa et Fujita, 1985, P. dissimilis kumejimaensis Ichikawa et Fujita, 1985, P. dissimilis hayashii Fujita, 2009.
Prosopocoilus (Psalidoremus) pseudodissimilis Kurosawa, 1976 (Fig. 1D)
Body length: male 28-63 mm, female 25-36 mm
Distribution: Japan (Yaeyama Iss.)
Subspecies: This species does not have subspecies.
Prosopocoilus (Psalidoremus) motschulskii (Waterhouse, 1869) (Fig. 1E)
Body length: male 24-60 mm, female 25-36 mm
Distribution: Taiwan Is.
Subspecies: This species does not have subspecies.
And seven morphological traits in Table 2 coded by binary notation and run through the SPSS hierarchical cluster analysis method and results are displayed in dendrogram in Fig. 2. In this analysis, common ancestor divided into two clades from inclinatus-hachijoensis cluster and others divided for later also divided into dissimilis and motschulskii - pseudodissimlis cluster and for the last motschulskii and pseudodissimlis divided in each area. But morphological characters not strong enough to assume speciation history of this subgenus, so geographical assessment also performed to provide strong hypothesis.
By considering ecological niche of the family Lucanidae, stag beetles not able to move across the sea surface. This provide strong barrier to the gene flow, which enables to proceed to speciation process by allopatric speciation. Also bathymetric elevation maps of distribution range at present sea level in Fig. 3, –750 m under sea surface in Fig. 4, –1,200 m under sea surface were displayed in Fig. 5. This provides strong evidence of the allopatric speciation of the subgenus Psalidoremus. Because of border of species almost identical to the land isolation gap appear on the bathymetric elevation maps in Figs. 3Fig. 4-5.
After all, subgenus Psalidoremus is allopatric group which sharing common ancestor in monophyletic clade. And their speciation scenario could be assumed by referring those morphological characters and bathymetric elevation and result as follows.
Hypothetical speciation scenario
First, common ancestor divided into inclinatus group and dissimilis group by Tokara Gap (marked as B in Fig. 6). Second, dissimilis group divided into two groups by dissimilis and motschulskii - pseudodissimilis cluster by Kerama Gap (marked as C in Fig. 6). Third, inclinatus - hachijo cluster and motschuskii - pseudodissimilis divided into each species level respectively by Hachijo Gap (marked as A in Fig. 6), and Taiwan Gap (marked as D in Fig. 6). This scenario has been pictorialized in Fig. 6 to enhance visibility of concept.
Discussion
This research has been performed to establish hyphothesis of how Psalidoremus species gone through the speciation from their common ancestors. And this is first attempt to provide visible clearance between Psalidoremus species and geographical data. Also Pictorial speciation scenario provides good visual agenda for other researchers.
The hypothesis in this here are supported by following references especially Ota (1998) which studied geographic pattern of amphibian and reptiles of Ryukyu archipelago has become good foundation of this article, and other taxonomic articles such as Fujita (2010) and Tsuchiya (2010), Tsuchiya (2017) provided numerous figures which in real sized magnification. Also, Tojo et al. (2017) provides strong evidence of deep straits provides similar difference in damselfly group, Repidolestis spp. Near Okinawa and Taiwan region.
Also this hypothesis could be disturbed by dispersal by typhoon as described in Osozawa et al. (2021) in case of Cicada species which distributes in Japan. Transportation by typhoon model was not considered in this article. Because of Lucanidae in general is not attempt to fly in raining day and also not good fliers as well.
In the result, most significant part is that the Hachijo gap is deep in substance, but the difference between P. inclinatus and P. hachijoensis not significant, also exposed in dendrogram. However, by consider in subspecies level, most closest relative is P. inclinatus miyakejimaensis which provides tendencies of intermediate tendencies in gradually from main island of Japan to Hachijo island. These tendencies should be study in advance in further researches. Also in subspecies level study must be performed to provide tendencies infraspecific level. Especially author noticed strong tendencies infraspecific difference in P. dissimilis, This tendencies would be the next research subject.
The scenario has been established in this research limited by the restriction of the morphological traits that useful in morphological identification and geographical patterns of each species. Therefore, research including DNA of each species and subspecies is demanded for more robust foundation of evidence.
Acknowledgments
This work was supported by National Research Foundation of Korea (NRF) grant funded by the Korea government (2013M3A9A5047052 and 2017M3A9A48999). Also author presenting great appreciation to those who cooperated in this research especially Jun-Tsong Lin, Associate Curator of National Taiwan Museum. And also Mr. Yamada sent important specimen from Japan.
References
GBIF Secretariat (2021, Retrieved August 30, 2021) GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus dissimilis Boileau, 1898 GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus dissimilis Boileau, 1898, from https://www.gbif.org/species/7867261
GBIF Secretariat (2021, Retrieved August 30, 2021) GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus hachijoensis Nomura, 1960 GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus hachijoensis Nomura, 1960, from https://www.gbif.org/species/7842151
GBIF Secretariat (2021, Retrieved August 30, 2021) GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus inclinatus (Motschulsky, 1857) GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus inclinatus (Motschulsky, 1857), from https://www.gbif.org/species/7487564
GBIF Secretariat (2021, Retrieved August 30, 2021) GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus motschulskii (Waterhouse, 1869) GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus inclinatus (Motschulsky, 1857), from https://www.gbif.org/species/6132326
GBIF Secretariat (2021, Retrieved August 30, 2021) GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus pseudodissimilis Kurosawa, 1976 GBIF Backbone Taxonomy. Checklist dataset. Prosopocoilus pseudodissimilis Kurosawa, 1976, from https://www.gbif.org/species/8787259
Figures and Tables
Fig. 1
Geographical patterns of the subgenus Psalidoremus. (A) Prosopocoilus inclinatus; (B) Prosopocoilus hachijoensis; (C) Prosopocoilus dissimilis; (D) Prosopocoilus pseudodissimilis; (E) Prosopocoilus motschulskii.
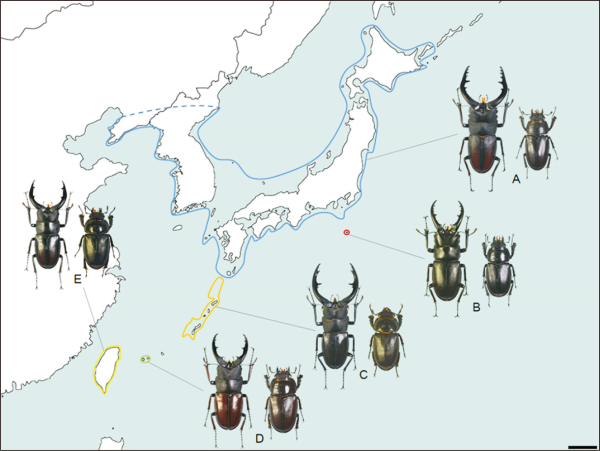
Fig. 2
Dendrogram of hierarchical cluster analysis in SPSS (ver. 28.0; IBM Co., Armonk, NY, USA) between species based on seven morphological traits. x-axis value means rescaled distance cluster combine.
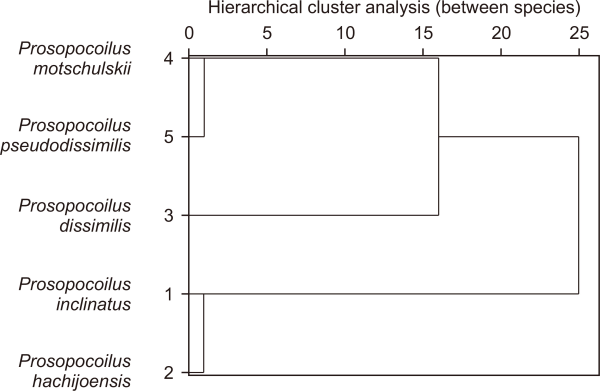
Fig. 3
Sea elevation map of present sea level. A-D indicates border of each species (Scale bar: 200 km).
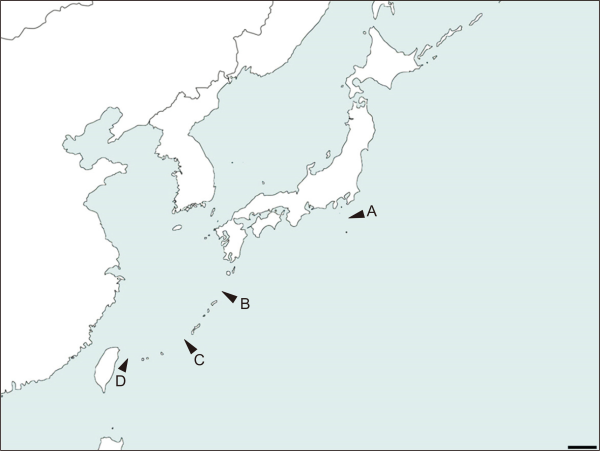
Fig. 4
Sea elevation map of sea level –750 m below surface. A-D indicates border of each species (Scale bar: 200 km).
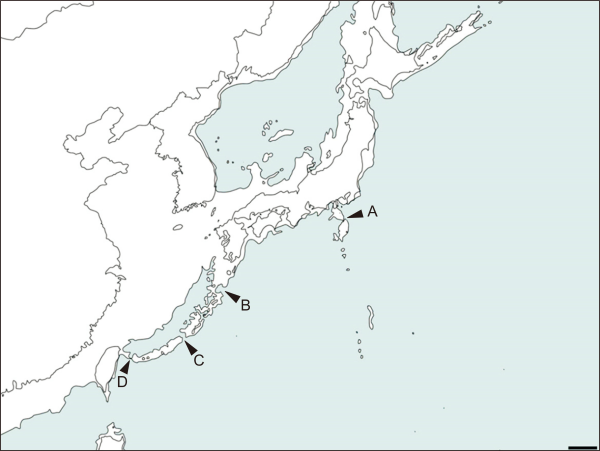
Fig. 5
Sea elevation map of sea level –1,200 m below surface. A-D indicates border of each species (Scale bar: 200 km).
Fig. 6
Pictorial speciation scenario of the subgenus Paslidoremus by each geographical gap A-D and speciation point provided in each gap marked with asterisk * (Scale bar: 200 km).
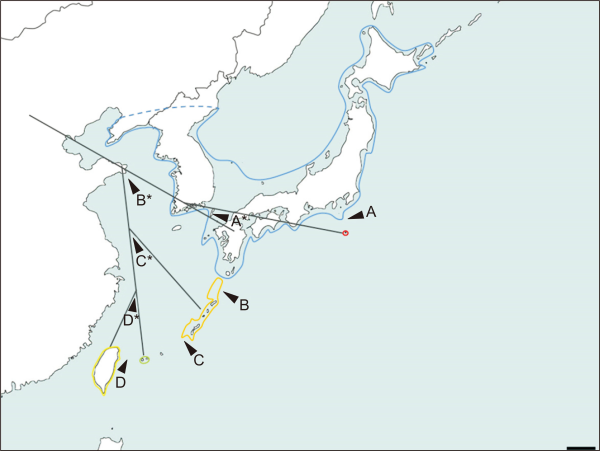
Table 1
Psalidoremus species analyzed in this research
| Species | Range | Possible closest relatives |
|---|---|---|
| Prosopocoilus inclinatus Motschusky, 1857 | Korea (Korean Peninsula, Uleung Is.), Japan (Honshu, Kyushu, Shikoku, Hokkaido, Izu Iss., Yakushima Is., Tanegashima Is., Mishima Is., Io Is., Kuroshima Is., Kuchinoerabu Is.), Russia (kunashir Is.), China (Liaoning Province) | P. hachijoensis |
| Prosopocoilus hachijoensis Nomura, 1960 | Japan (Hachijo Is.) | P. inclinatus |
| Prosopocoilus dissimilis Boileau, 1898 | Japan (Tokara Iss., Amami Iss., Okinawa Is.) | P. inclinatus |
| Prosopocoilus pseudodissimilis Kurosawa, 1976 | Japan (Iriomote Is., Ishigaki Is.) | P. motschulskii |
| Prosopocoilus motschulskii Waterhouse, 1869 | Taiwan | P. pseudodissimilis |