Introduction
The environment in which organisms live is always changing, and the effect of these changes on the organisms within is highly complex (Nagelkerken & Munday, 2016; Stephens, 1991). In order to clearly understand the effects that environmental changes have on organism, it is therefore necessary to conduct long-term monitoring and analysis so that trends are revealed (Frommolt and Tauchert, 2014; Hashimoto et al., 2020). Environmental factors, including temperature, altitude, precipitation, and solar radiation, are known to affect the habitats of wildlife, including bird species (Martin & Wiebe, 2004; Salewski et al., 2013). In general, birds show various behaviors and preference when choosing their habitat (Fisher & Davis, 2010; Maxwell et al., 2018). For example, forest birds are affected by their physical environment in terms of temperature and rainfall (Cormont et al., 2011; Grinde et al., 2017). Thus, wetland conditions, regarding habitats for birds, are characterized by the structure of the wetland, water quality, provision of food and hiding sites, and the presence of predators (Whited et al., 2000; Yuan et al., 2014). Wetlands also provide productive habitats for aquatic and terrestrial birds as well as wintering grounds for migrating birds (VanRees-Siewert & Dinsmore, 1996).
Many surveys on wetlands and their surrounding areas (Ismail et al., 2012; Park, 1990; Rathod et al., 2016; Sharma et al., 2014) were conducted to better understand the diversity and structure of bird communities, which is necessary to determine the importance of regional or local landscapes for avian conservation (Kattan & Franco, 2004). Changes in species richness, evenness and individual numbers may be used to compare current ecological conditions to previous ones, and they can also be used to predict future ecological conditions (Kim, 2008).
Long-term monitoring studies have been conducted in Jeju Island (Banjade et al., 2019; Park, 1990) but only for some areas, and long-term monitoring data is required for other areas where research has not yet been conducted. Various studies have been conducted in Hallasan National Park, and these have focused on bird distribution, habitat information, and population structure (Kim et al., 2014; Koh & Kim, 2011). However, the impact that various environmental factors can have not yet been studied.
Hence, we investigate here the relationship between the environmental factors, such as precipitation, temperature and seasons, upon the bird species diversity, richness and evenness in the Mulchat-oream wetlands in Hallasan National Park.
Materials|Methods
Study area
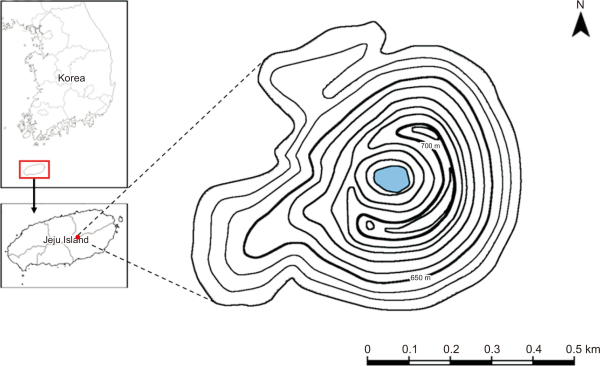
The study was conducted at the Mulchat-oream wetlands (E126° 40′ 12.9, N33° 31′ 11.4, alt. 670 m). These are present in a volcanic mountain containing a freshwater Crater Lake at its summit (Fig. 1). This wetland area is protected by law under the Ministry of Environment since it is rich in biodiversity and maintains its primitive nature. In the past, people were permitted to enter, but the natural ecosystem was damaged by human presence and visitors have since been banned. The study site contains diverse flora and fauna, and the plant species are composed of Carpinus laxiflora Siebold & Zucc, Styrax japonicus Siebold & Zucc, Quercus serrata Murray in the upper part and Sasa quelpaertensis Nakai in the lower part. The faunal diversity includes Capreolus pygargus, Cervus nippon, Sus scrofa and occasional sighting of Cervus elaphus. The International Union for Conservation of Nature red list birds such as Pitta nympha and Terpsiphone atrocaudata have been frequently observed in the summer.
Data collection
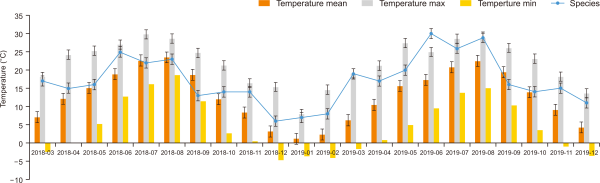
The field study was carried out from March 2018 to December 2019. Data were collected seasonally in each surveyed year, and on the clear and sunny days, the survey was conducted between 06.00-10.00 hr and from 14.00-18.00 hr to avoid bias caused by the weather conditions. Birds were observed using Olympus binoculars (10×50 DPSI; Olympus, Tokyo, Japan) and were identified to the species level by calls and excrement remnants by using the standard reference book Lee et al. (2020). Species were categorized following the reference of the Ornithological Society of Korea (2009) and the National Species List of Korea (National Institute of Biological Resources, 2020). HOBO RX3000-Remote monitoring and a weather station (Onset, Bourne, MA, USA) was set and identified the mean monthly fluctuation on the temperature and precipitation (Fig. 2). During the study period, the data were collected by linking to the HOBO site once a month with a measurement interval of 60 minutes. The average winter temperature during the study period was 1.42°C and the summer temperature was 20.94°C. Seasonal precipitation varied from 238 mm in winter and 977 mm in summer.
Data analysis
The data were analyzed using a linear analysis. All other procedures were implemented using Excel 2018 (Microsoft, Redmond, WA, USA) and SPSS 20.0 (IBM Co., Armonk, NY, USA). The population index was calculated using the following formula:
∘ Dominance index (Brower et al., 1990) : DI : DI=ni/N DI : dominance index, N : total individual, ni : individual of the ith species
∘ Biodiversity index (Shannon & Weaver, 1949) : DH'=-∑{(ni/N) ․ ln(ni/N)} (ni: species of I’ individual, N: total individual)
∘ Evenness index (Margalef, 1994) : E’ E’=D’/Ln (S), E’=evenness, D’=diversity, S: total species
∘ Richness index : R’ R’=(S-1/Ln(n), R’=richness, S: total species, N: total individual
Results
Aspect of the entire bird population
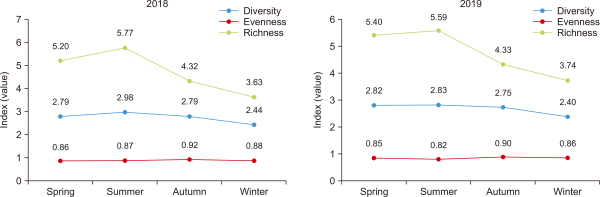
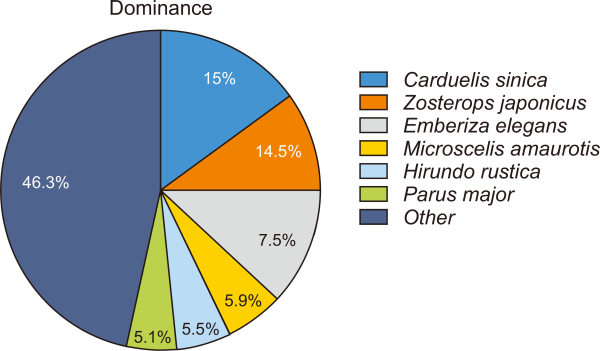
A total of 255 individuals of 37 species were recorded in this study. In a 2018 survey, 211 individuals belonging to 36 species were recorded, and in 2019, 242 individuals belonging to 37 species were recorded (Supplementary Table 1). Furthermore, the species diversity was 2.44 to 2.79, species richness was 3.63 to 5.77 and species evenness was 0.86 to 0.92 in 2018. The species diversity was 2.40 to 2.83, species richness was 3.74 to 5.59 and species evenness was 0.82 to 0.90 in 2019 (Fig. 2). The highly dominant species are Carduelis sinica (15%) followed by Zosterops japonicus (14.50%), Emberiza elegans (7.50%), Microscelis amaurotis (5.90%), Hirundo rustica (5.50%), and Parus major (5.10%) (Fig. 3).
Comparison of the population with weather factors
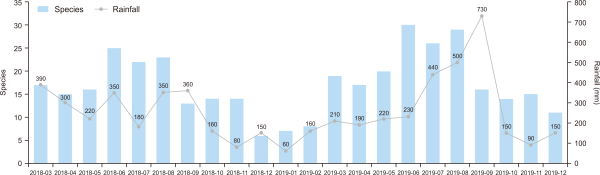
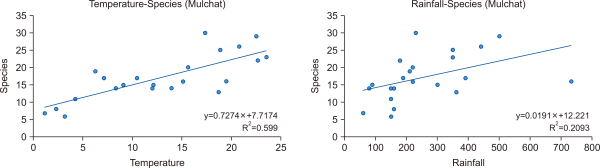
The average seasonal temperature and precipitation in the Mulchat-oream wetlands compared to the species presence is shown in Figs. 4 and 5. The appearance of the birds was lowest in the winter and highest during the summer (Table 1). The analysis showed that the species richness and evenness increased with an increase in temperature and precipitation (Fig. 6). However, the number of species appearing in the Mulchat-oream wetlands is significantly dependent on the temperature (P<0.001, df=2). One notable thing about the precipitation was that the precipitation value in the autumn was higher than the precipitation value in the summer by the LINGLING and TAPAH typhoon.
Discussion
The opening of forest roads and expansion of trails tends to greatly reduce bird species diversity because trees are eliminated in the area, resulting in simplification of the forest environment (Aleixo, 1999; Thiollay, 1997; 1999). Also, frequent climbing activities can reduce bird population density due to constant interference (Camp & Knight 1998; Covy et al., 2019). Mulchat-oream is restricted to visitors, but travelers usually visit because of the surrounding forest and its connection with the Saryeoni forest trail (Seok et al., 2010). The bird species diversity at Mulchat-oream is generally low compared to that at the Hallasan National Park (Oh, 2002; Park et al., 2011). The occurrence of a lower number of species at the Mulchat-oream wetlands could be attributed to frequent human interference. However, species that prefer the edges of forests or the interior of forests are different, making it difficult to say that human interference has hindered bird inhabitation (Baker et al., 2002; Laiolo & Rolando, 2006). Therefore, it is believed that further investigation is needed to detail the habitat preferences and to conduct bird surveys on the trails (Bell, 1996; Deluca & King, 2014).
We have observed a higher number of Passeriformes in this study at the site for both surveyed years. The presence of a higher number of Passeriformes is related to the absence of aquatic birds (Moreno-Mateos et al., 2009; Rosselli & Stiles, 2012). The absence of diverse submerged plants, deep water depth and isolated geographical locations makes difficult for aquatic birds to prefer this area, and this has resulted in the dominance of Passeriformes (Pöysä et al., 2013).
The seasonally analyzed species diversity and evenness showed a relatively stable population of 2.40 to 2.90 and 0.82 to 0.90, respectively, but the richness showed seasonal variations of 3.63 in the winter to 5.70 in summer. In general, the species diversity, evenness, and richness are influenced by geographical, meteorological, physical, and biological factors (Mäkeläinen & Lehikoinen, 2021). Jeju Island is a geopolitical stopover site for migratory birds, especially in the middle of Mt. Hallasan Mountain, where summer visitor birds breed and spend their summer time (Kim et al., 2010; 2019). Human interference during the breeding season is limited since the area is restricted for visitors. Therefore, it is expected to have a higher species richness during the summer compared to any other seasons. However, studies on food resources, such as invertebrates and insects, have not yet been conducted, so it is hard to predict the relationship among the species richness and other biological factors. Furthermore, various factors need to be investigated that affect the bird populations.
To a large extent, the climatic preferences of the species determine their response to climate change (Nichelmann & Tzschentke, 2002). Temperature preferences play a key role in driving trends in animal populations (Gaüzère et al., 2016). However, the relative importance of the temperature and precipitation preferences is still poorly understood, particularly in systems where ecological processes are strongly constrained by the amount and timing of rainfall (Knape & de Valpine, 2010). In this study, we have estimated the role played by the temperature and precipitation preferences in determining the population trends for birds in mountain wetlands. Trends were derived through biodiversity monitoring data and temperature and precipitation preferences were estimated from species appearance data. The results showed that the temperature trends were first and that precipitation is also related to the birds. A comparison of these shows that the precipitation preferences are less significant compared to temperature. Climatic factors, such as temperature and precipitation, may affect the bird communities directly via physiological constraints, for instance by restricting the activity, mobility and foraging time of the birds (Filloy et al., 2015).
The Korean Peninsula has historically offered a temperate climate (Lim et al., 2018), but it has recently been changing to a subtropical climate (Lee et al., 2017) due to global climate change (Wuebbles & Jain, 2001). These climatic factors create confusion upon migratory and native birds species that directly influences reproductive success. Climate change is causing mismatches in peak food availability, snow cover and other factors that could severely impact successful migration and reproduction of avian populations unless they are able to adjust to new conditions (Jetz et al., 2007). Resident birds also face challenges if precipitation and/or temperature patterns vary in ways that result in a mismatch for food and breeding (Rodenhouse et al., 2008).
Thus, this study shows the environmental factors and seasonal patterns that affect bird populations. It is believed that if long-term monitoring is carried out, the impact that temperature and precipitation have upon bird populations will be more clearly established.
Figures and Table

The location of study site in Mt. Halla Mulchat-oream on Jeju island.

Bird diversity value at each season in the study region.

Dominant species by observation on the study site.

Comparison of temperature with number of species of birds for four seasons over two years.

Comparison of precipitation with number of species of birds for four seasons over two years.

The correlation between rainfall and temperature with number of species on Mulchat-oream.

Liner regression analysis data refereeing to temperature and precipitation